by Paul K. Mensah, Carolyn G. Palmer and Oghenekaro N. Odume
http://dx.doi.org/10.5772/60767
© 2015 The Author(s). Licensee InTech. This chapter is distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
NOTICE: THIS WORK MAY BE PROTECTED BY COPYRIGHT
YOU ARE REQUIRED TO READ THE COPYRIGHT NOTICE AT THIS LINK BEFORE YOU READ THE FOLLOWING WORK, THAT IS AVAILABLE SOLELY FOR PRIVATE STUDY, SCHOLARSHIP OR RESEARCH PURSUANT TO 17 U.S.C. SECTION 107 AND 108. IN THE EVENT THAT THE LIBRARY DETERMINES THAT UNLAWFUL COPYING OF THIS WORK HAS OCCURRED, THE LIBRARY HAS THE RIGHT TO BLOCK THE I.P. ADDRESS AT WHICH THE UNLAWFUL COPYING APPEARED TO HAVE OCCURRED. THANK YOU FOR RESPECTING THE RIGHTS OF COPYRIGHT OWNERS.
Abstract
The use of agrochemicals, especially herbicides, is necessary to control pests in order to produce adequate food for the global population (estimated at 7 billion). Glyphosate and glyphosate-based herbicides have been used extensively for this purpose but recent studies have reported these chemical substances to be found in aquatic ecosystems, wildlife and humans in various quantities. In this chapter, we reviewed the impacts of glyphosate and glyphosate-based herbicides on wildlife and humans using measured endpoint effects caused by genotoxicity, cytotoxicity and reproductive toxicity. We used findings from different current investigations to demonstrate adverse effects, or otherwise, of glyphosate exposure to wildlife and humans. Our review reveals that glyphosate and its formulations may not only be considered as having genotoxic, cytotoxic or endocrine disrupting properties but they may also be causative agents of reproduction abnormalities in both wildlife and humans. Furthermore, the extensive use of glyphosate-based herbicides in genetically modified glyphosate-resistant plants grown for food and feed should be of grave concern since they can be sources of genotoxicity, cytotoxicity, and reproductive toxicity in wildlife and humans.
Keywords: Cytotoxicity, genotoxicity, glyphosate, human toxicity, wildlife toxicity
1. Introduction
The use of agrochemicals is necessary to control pests and increase yields in order to produce adequate food for the global population, estimated at 6.8 billion in 2009 [1], and recently reported to have reached 7 billion [2]. Developing countries, where 1.02 billion people (15 %) are undernourished and 1.3 billion people (19 %) live on an inadequate diet [1], need an adequate food supply. However, the agricultural sector’s annual application of over 140 billion kilograms of fertilizers and large amounts of pesticides creates massive sources of diffuse pollution of freshwater ecosystems [3]. In an attempt to increase food production, there is extensive use of herbicides without much regard to the consequences posed to the environment and humans. Glyphosate-based herbicides, which are extensively used in genetically modified glyphosate-resistant plants, are found all over the world [4] and have been reported to occur in various quantities in the aquatic ecosystem, wildlife and humans.
Globally, the presence of pesticides accumulation in both wildlife and humans is on ascendancy, with the health and normal functioning of the endocrine systems being at risk [5-7]. It is believed that the effects of these chemicals on normal functioning of the endocrine system are responsible for a number of developmental anomalies in a wide range of species, from invertebrates to higher mammals [8-11].
The aquatic environment is a receptacle of several undesirable contaminants, including agrochemicals. Therefore, contamination of the aquatic environment by pesticides has become a huge environmental concern worldwide [12]. Glyphosate and glyphosate-based herbicides are among the most widely used class of pesticides. Roundup®, is a major glyphosate-based herbicide used worldwide. Over the years, studies have suggested adverse effects of glyphosate and glyphosate-based compounds on terrestrial and aquatic environments, but recent publications are alluding to the possible effects of glyphosate on mammals, including humans, at different levels of biological organisations as well. In this chapter, the toxicology of glyphosate and glyphosate-based herbicides are explored. We reviewed the impacts of glyphosate and glyphosate-based herbicides on wildlife and humans using measured endpoint effects caused by genotoxicity, cytotoxicity and reproductive toxicity.
1.1. Pesticide as pollutants of freshwater ecosystems
Pesticides are mixtures of chemical substances designed to control, repel, mitigate, kill or regulate the growth of undesirable and nuisance biological organisms [13]. Pests include plant pathogens, weeds, nematodes, molluscs, insects, fish, birds, mammals and microorganisms such as bacteria and viruses. They compete with humans for food, transmit diseases and destroy crops as well as properties [13]. There are various ways of classifying pesticides, with the classification based on the type of pest they control being the most common. For example, insecticides, herbicides, fungicides, nematicides and rodenticides are used to control insects, weeds, fungi, nematodes and rodents, respectively. Furthermore, majority of pesticides are synthetic as they are formulated through industrial processes, while a few are biological as they are derived from natural sources. In addition, broad-spectrum pesticides are applied in controlling a wide range of species but narrow-spectrum pesticides control a small group of pests [13].
Although pesticides are used in agriculture to maintain high production efficiency, they may be environmental hazards and pose risk particularly to non-targeted organisms, and generally to aquatic ecosystems [14, 15]. The potential of a pesticide’s risk to an aquatic ecosystem is influenced by its properties, including half-life, mobility and solubility [13]. Microbial activity, drainage pattern, rainfall, treatment surface and application rate can also affect pesticidal activity on a local, regional or global scale [16, 17]. Pesticides get into aquatic systems through processes such as direct applications, surface runoffs, spray drifts, agricultural returns and groundwater intrusions [18]. Pesticides found in urban and agricultural settings in recent times have been implicated in the deaths of many aquatic biota [19].
1.2. Presence of herbicides in freshwater ecosystems
Weeds are unwanted vegetation, which are not planted intentionally, but inadvertently grow in unexpected places. They are usually controlled (i.e., killed or suppressed) by the application of a specific herbicide type or class. Classification of herbicides may depend on the criteria used. The two most common criteria employed in herbicide classification are based on time of application and mode of action [20]. Table 1 shows herbicides classification based on time of application and mode of action. Herbicides, which are widely used to control weeds in forestry and agriculture, can reach the aquatic ecosystems by uncontrolled runoff, aerial drift or inadvertent overspray. In some cases, herbicides are directly sprayed at aquatic weeds (e.g. water hyacinth) found on surfaces of water bodies as a control measure. All these impact the aquatic biota.
Classification / Chemical family* / Examples
Time of herbicide application / -- / --
Pre-emergence: applied to the soil after the crop is planted / Dinitroaniline / Pendulum AquaCap Oryzalin (Surflan AS)
Post-emergence: applied to both crop and weeds after they have germinated and emerged from the soil / Benzoxazole / Acclaim® Extra
Mode of action / -- / --
Hormone inhibitors: These herbicides inhibit cell division and growth in the meristem regions (growing points) by mimicking IAA, the natural plant hormone. This interferes with cell wall plasticity and nucleic acid metabolism. / Phenoxycarboxylic acid / 2,4-D; 2,4-DB; 2,4-DP; MCPA; MCPB
Cell division inhibitors: These herbicides bind to tubulin, the major microtubule protein, to form a herbicide–tubulin complex / Carbamate / Chlorpropham; Propham; Carbetamide
leading to a loss of microtubule structure and function. Herbicide-induced microtubule loss may cause cells to neither divide nor elongate, which may be observed as swelling of root tips. / -- / --
Photosynthesis inhibitors: These herbicides inhibit photosynthesis by preventing electron flow, CO2 fixation and, ATP and NADPH2 production in the photosystem II complex in chloroplasts. Lack of ATP and NADPH2, as well as free radicals destroy cell membranes lead to eventual death of plant. / Triazine / Atrazine, Simazine, Caparol
Lipid synthesis inhibitors: These herbicides inhibit fatty acid and lipid biosynthesis. This causes reduction in cuticular wax development and eventual death of plant. / Thiocarbamate / Cycloate; Dimepiperate; Pebulate; Thiobencarb; Triallate
Cell metabolism inhibitors: These herbicides capture electrons from photosystem I, reduce them to form herbicide free radicals, which then destroy cell membranes. / Bipyridylium / Diquat; Paraquat; Gramoxone
EPSP Synthase Inhibitors: These herbicides inhibit EPSP synthase enzyme, which leads to the depletion of the aromatic amino acids tryptophan, tyrosine and phenylalanine. / Glycines / Glyphosate
Table 1. Examples of herbicide classification based on time of application and mode of action (* Note: there may be more than one chemical family for each category of herbicide)
The potential of some herbicides to control unwanted vegetation is inherent in their chemical nature, while others have additives to enhance their efficacy. These additives include carriers and adjuvants. In recent years, carriers and adjuvants have been implicated in adding to the toxicity of the active ingredients, and in some cases, have been even more toxic than the active ingredient alone [20]. Prior to the registration of herbicide products for use, not only does the herbicidal properties (Table 2) are assessed, but also the potential effects on humans, animals and environmental safety are assessed. The inherent toxicity of a herbicide, concentration to which an organism is exposed, and duration of exposure determine the extent to which the herbicide can adversely affect an aquatic organism [21]. Herbicides may reach aquatic ecosystems directly by an overhead spray of aquatic weeds, or indirectly through processes such as agricultural runoff, spray drift and leaching [13]. Potential problems associated with herbicide-use include injury to non-target vegetation, injury to crops, residue in soil or water, toxicity to non-target organisms, and concerns for human health and safety [20]. Herbicides can influence the environmental water quality and ecosystem functioning by reducing species diversity, changing community structure, modifying food chains, altering patterns of energy flow and nutrient recycling, as well as reducing resilience of ecosystems [22].
Herbicidal property / Explanation
Chemical structure / The biologically active portion of a herbicide product is the active ingredient. It is the fundamental molecular composition and configuration of the herbicide. The physical and chemical properties of a herbicide can also determine the method of application and use.
Water solubility and polarity / Herbicides that are produced as salts dissolve quite well in water and are usually formulated to be applied in water, while non-polar herbicide sources are not. Water is the main substance used to disperse (spray) herbicides, and hence the water solubility of a herbicide influences the type of product that is formulated, how it is applied and the movement of the herbicide in the soil profile.
Volatility / Herbicides with a high vapour pressure volatilise easily, while those with a low vapour pressure are relatively non-volatile. The volatility of a herbicide can determine the mode of action and the herbicide’s fate in the environment.
Formulations / Commercial herbicide products contain an active ingredient and “inert” ingredients. An “inert” ingredient could be a carrier that is used to dilute and disperse the herbicide (e.g. water, oil, certain types of clay, vermiculite, plant residues, starch polymers, certain dry fertilizers) or an adjuvant (e.g. activator, additive, dispersing agent, emulsifier, spreader, sticker, surfactant, thickener, wetting agent) that enhances the herbicide’s performance, handling, or application.
Table 2. Herbicidal properties of herbicides that enhance their efficacy
2. Glyphosate and glyphosate-based herbicides
Glyphosate (N-(phosphonomethyl) glycine) (Figure 1) and glyphosate-based herbicides are the world’s leading post-emergent, organophosphonate systemic, broad-spectrum and nonselective herbicides for the control of annual and perennial weeds [22, 23]. Worldwide, the number one glyphosate-based herbicide used is Roundup®. Other trade names of glyphosatebased herbicides include Roundup Ultra®, Roundup Pro®, Accord®, Honcho®, Pondmaster®, Protocol®, Rascal®, Expedite®, Ranger®, Bronco®, Campain®, Landmaster®, Fallow Master® and Aquamaster® manufactured by Monsanto; Glyphomax®, Glypro® and Rodeo® manufactured by Dow Agrosciences; Glyphosate herbicide manufactured by Du Pont; Silhouette® manufactured by Cenex/Land O’Lakes; Rattler® manufactured by Helena; MirageR® manufactured by Platte; JuryR® manufactured by Riverside/Terra; and Touchdown® manufactured by Zeneca [24-26].
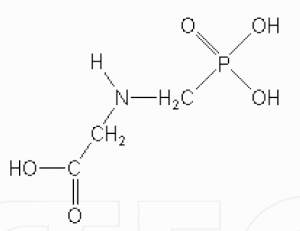
Figure 1. Molecular structure of N-(phosphonomethyl) glycine
Glyphosate has relatively low solubility in water (12 g/L at 25° C and 60 g/L at 100° C), but is insoluble in other solvents [27]. Therefore, commercial formulations of glyphosate are usually in the form of salt to ensure higher solubility yet maintaining the herbicidal properties of the parent compound [22]. Formulations of glyphosate in salt form include monoammonium salt, diammonium salt, isopropylamine salt, potassium salt, sodium salt, and trimethylsulfonium or trimesium salt. Of these, the isopropylamine, sodium, and monoammonium salt forms are commonly used in formulated herbicide products [28]. The isopropylamine salt is the most commonly used in commercial formulated products (e.g. Roundup®). The concentration of glyphosate is commonly expressed as mg a.i./L (active ingredient/Litre) or mg a.e./L (acid equivalents/Litre) [22]. Acid equivalent is the theoretical per cent yield of parent acid from a pesticide active ingredient, which has been formulated as a derivative (usually esters, salts or amines) [29].
2.1. Mode of action of glyphosate
As a systemic herbicide, glyphosate is readily translocated through the phloem to all parts of the plant. Glyphosate molecules are absorbed from the leaf surface into plant cells where they are symplastically translocated to the meristems of growing plants [22]. Glyphosate’s phytotoxic symptoms usually start gradually, becoming visible within two to four days in most annual weeds, but may not occur until after seven days in most perennial weeds. Physical phytotoxic symptoms include progress from gradual wilting and chlorosis, to complete browning, total deterioration and finally, death [22]. The primary mode of action of glyphosate is confined to the shikimate pathway aromatic amino acid biosynthesis, a pathway that links primary and secondary metabolisms.
Shikimate (shikimic acid) is an important biochemical intermediary in plants and microorganisms, such as bacteria and fungi. It is a precursor for the aromatic amino acids phenylalanine, tryptophan and tyrosine. Other precursors of the shikimate pathway are indole, indole derivatives (e.g. indole acetic acid), tannins, flavonoids, lignin, many alkaloids, and other aromatic metabolites. The biosynthesis of these essential substances is promoted by enzyme 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS), the target enzyme of glyphosate. This enzyme is one of the seven enzymes that catalyse a series of reactions, which begins with the reaction between shikimate-3-phosphate (S3P) and phosphoenolpyruvate (PEP). The shikimate pathway accounts for about 35 % of the plant mass in dry weight and therefore any interference in the pathway is highly detrimental to the plant. Glyphosate inhibits the activity of EPSPS, preventing the production of chorismate – the last common precursor in the biosynthesis of numerous aromatic compounds in bacteria, fungi and plants. This causes a deficiency in the production of the essential substances needed by the organisms to survive and propagate [22, 30]. The pathway is absent in animals, which may account for the low toxicity of glyphosate to animals.
However, acute effects in animals, following intraperitoneal administration of high glyphosate doses, suggest altered mitochondrial activity, possibly due to uncoupling of oxidative phosphorylation during cellular respiration [27]. In summary, glyphosate ultimately interrupts various biochemical processes, including nucleic acid synthesis, protein synthesis, photosynthesis and respiration, which are essential life processes of living things.
2.2. Environmental fate of glyphosate
Glyphosate has a strong soil adsorption capacity, which limits its movement in the environment. The average half-life of glyphosate in soil is two months, but can range from weeks to years [24]. Glyphosate in freshwater ecosystems has an average half-life of two to ten weeks [24]. The rate of degradation in water is generally slower than in most soils because of fewer microorganisms in water than in soils [31]. When glyphosate undergoes degradation, it produces aminomethylphosphonic acid (AMPA) and carbon dioxide [32], both of which reduce pH when dissolved in water. However, pH is known to affect the stability of glyphosate in water. For instance, glyphosate did not undergo hydrolysis in buffered solution with a pH of 3, 6 or 9 at 35° C, while insignificant photodegradation has been recorded under natural light in pH 5, 7 and 9 buffered solutions [28]. In freshwater ecosystems, glyphosate dissipates through degradation, dilution, and adsorption on organic substances, inorganic clays and the sediment (the major sink for glyphosate in water bodies) [24, 31]. With its long half-life and its ability to cause death of organisms in aquatic ecosystems, it is recommended that glyphosate should be used as an aquatic herbicide to treat only one-third to half a water body at any one time [24].
2.3. Toxicology of glyphosate and its effects on aquatic organisms
In recent years, the exposure of non-target aquatic organisms to glyphosate-based herbicides has aroused great concern globally because of high water solubility and the extensive use of glyphosate-based herbicides [25]. In this regard, polyoxyethylene amine (POEA), a surfactant, has been implicated as being the main cause of the relatively high toxicity of Roundup® to several freshwater invertebrates and fishes [25, 33]. Technical grade glyphosate is slightly to very slightly toxic, with reported LC50 values of greater than 55 mg/L and a 21 d NOEC (no observed effect concentration) value of 100 mg/L [25, 33].
Conversely, formulations of glyphosate are moderately to very slightly toxic with 2 d EC50 values of 5.3-5600 mg/L and 21 d MATC values of 1.4-4.9 mg/L reported [27]. The LC50 values also determine which glyphosate formulation can be applied in aquatic ecosystems. It should be noted that high LC50 value of a chemical substance to an organism implies low toxicity of that particular chemical substance to that particular organism, and the reverse is also true. For instance, Rodeo® has relatively high LC50s (>900 mg/L) for aquatic species and is permitted for use in aquatic ecosystems, while Touchdown 4-LC® and Bronco® have low LC50 values for aquatic species (<13 mg/L), and are not registered for aquatic use [24]. Similarly, Roundup® is not registered for use in aquatic ecosystems in the United States because its 96 h LC50 value for Daphnia is 25.5 mg/L, while that of glyphosate alone is 962 mg/L [24].
In recent years, glyphosate has been found in surface waters long after it has been used to control aquatic weeds, although it is generally regarded as having a low potential for contaminating surface waters [13, 34]. In fact, its mode of action was designed to affect only plants [30], but various studies in recent years have reported adverse impact on nontarget animals [25, 33, 35].
2.4. Wildlife toxicology of glyphosate and glyphosate-based herbicides
Wildlife ecology is the application of ecological principles to the study of wildlife species. The term wildlife, however, lacks a universally accepted definition, and its common use changed during the 1900s in association with development of the profession of wildlife management. Historically, wildlife management focused on hunted or harvested birds and mammals that were collectively referred to as game species. However, current description of wildlife ecology includes different levels of biological organisation as well as individual organisms within a population and their interactions with the environment [36]. The effects of environmental contaminants on the health and persistence of wildlife populations have been a concern of environmentalists for many decades [36]. This led to increased interest in the study of exposure of wildlife to environmental contaminants, and hence wildlife toxicology.
A growing collection of wildlife toxicological studies examining diverse wildlife species demonstrates that exposure to environmental contaminants over the years is the cause of increasing disappearance of certain species. Such a threat to global biodiversity usually starts with developments of abnormalities in contaminated organisms. These may include the disruption of genetic material, cell integrity and major but subtle birth defects in individual species. This is against the belief, held for a long time since introduction of glyphosate as a commercial herbicide in the 1970s, that glyphosate is non-toxic to wildlife and humans. In fact, US EPA has classified glyphosate as a “Group E carcinogen”, implying it is “non-carcinogenicity for humans” [37]. The main reason for this has been attributed to the fact that the herbicidal activity of glyphosate targets specifically inhibition of the shikimate pathway, which is only present in plants and microorganisms, but conspicuously absent in animals and humans [38]. However, recent studies seem to suggest that glyphosate has genotoxicity, cytotoxicity and reproductive toxicity in wildlife and humans [39-40].
Genotoxicity is a term used to describe the destructive effect by toxic agents referred to as genotoxins on a cell's genetic material (DNA, RNA). Genotoxins include both radiation and chemical genotoxins. There are three primary effects that genotoxins can have on organisms by affecting their genetic information depending on the type of genotoxin. These include cancer-causing agents (carcinogens), mutation-causing agents (mutagens) or birth defect-causing agents (teratogens) [41]. Conversely, cytotoxicity is the term used to describe destructive effect of cells by agents referred to as cytotoxins. Cytotoxic cells may undergo necrosis, in which they lose membrane integrity and die rapidly as a result of cell lysis; they can stop growing and dividing; or they can activate a genetic program of controlled cell death, also referred to as apoptosis [41]. Reproductive toxicity refers to the toxic effects of a substance on the reproductive ability of an organism and the development of its offspring. The Globally Harmonized System (GHS) defines reproductive toxicity as adverse effects of chemical substances on sexual function and fertility in adult males and females, as well as developmental toxicity in the offspring [42]. Developmental toxicity refers to adverse effects induced during pregnancy, or as a result of parental exposure to a chemical substance manifested at any point in the life span of the organism [42]. Thus, reproductive toxicity may be grouped into two main categories: adverse effects on reproductive ability or capacity, and adverse effects on development of the offspring. Developmental toxicity and reproductive toxicity tests are usually performed with female and male animals, embryonic stem cells or whole embryos. In the following sections, we review a few of the many reported studies on adverse effects caused by glyphosate and glyphosate-based herbicides as a result of genotoxicity, cytotoxicity and reproductive toxicity in wildlife as well as in humans.
3. Toxicity of glyphosate and glyphosate-based herbicides to wildlife
It has been argued that the seasonality of crop cultivation and subsequent seasonal application of herbicides means that their presence in aquatic ecosystems is typically periodic. More so, episodic exposures are often for a short period, which is followed by long periods of nonapplication that has the capacity to “water down” the exposure effects. Therefore, the assessment of genotoxic endpoints in fish after removal of the contamination source is essential to understanding the dynamics of herbicide genotoxicity and risk posed by these agrochemicals. In order to test this assertion, Guilherme et al. [12] investigated the genotoxic potential of Roundup® to European eel (Anguilla anguilla) [12]. A. anguilla was exposed to 116 μg/L Roundup® for 3 days, and allowed to recover for 1, 7 and 14 days after exposure has ceased. A comet assay was applied to identify DNA damage in blood cells during both exposure and post-exposure periods. Guilherme et al. [12] reported that Roundup® has genotoxic potential and exhibited oxidative DNA damage (pyrimidine bases oxidation), although a recovery was evident when considering non-specific DNA damage on day 14 post exposure. Roundup® was able to induce a late oxidative DNA damage (day 14). In the report, Guilherme et al. suggested that blood cells of A. anguilla exposed to Roundup® appeared to be more successful in repairing damage with a non-specific cause than that associated with base oxidation [12]. Overall, the findings of this study reinforce the hypothesis that Roundup® as an agrochemical may cause long-lasting damage to fish due to genotoxicity.
In another study, Vera-Candioti et al. evaluated the genotoxicity and cytotoxicity of glyphosate- based formulations Panzer and Credit® after exposure to Cnesterodon decemmaculatus (Jenyns, 1842) (Pisces, Poeciliidae) under laboratory conditions [43]. They used micronuclei induction (MN) and alterations in the erythrocytes to erythroblasts ratio for genotoxicity and cytotoxicity as endpoints, respectively. They reported that both 3.9 and 7.8 mg/L of Panzer increased MN frequency at 48 and 96 h of treatment. Similarly, C. decemmaculatus exposed to tested concentration of Credit® had increased frequency of MN over control group after 96 h, but not after 48 h. However, they reported that both Panzer and Credit® treatment did not cause cytotoxicity in fish, regardless of the concentration and sampling time. In their conclusion, Vera-Candioti et al. suggested that Panzer and Credit® should be considered as glyphosate- based commercial formulations with genotoxic but not cytotoxic effect properties [43].
Low levels of glyphosate-based herbicide have also been reported to cause adverse effects on reproduction (fecundity) and clutch size of aquatic invertebrates. In a study, Cuhra et al. reported that Daphnia magna exposed to 0.45 mg/L Roundup® significantly decreased fecundity compared to the control group [44]. They also reported that animals exposed to 1.35 mg/L Roundup® reached reproductive age, but almost all eggs and developing embryos were aborted and did not hatch, whereas animals exposed to 4.05 mg/L Roundup® died before reaching maturation. However, fecundity in Daphnia magna exposed to Roundup® concentrations of 0.05 and 0.15 mg/L was not significantly affected. The abortion rates for animals exposed to Roundup® concentrations of 0.05, 0.15 and 0.45 mg/L were not significantly different from those of the control group. On the other hand, the abortion rates for animals exposed to 1.35 mg/L of Roundup® were significantly higher than control group, reaching nearly 100 %. Cuhra et al. also reported that the size of first clutch (brood) juveniles born from groups exposed to 0.05, 0.15 and 0.45 mg/L Roundup® were not significantly different from the control group [44]. However the size of second clutch juveniles from animals exposed to 0.05, 0.15 and 0.45 mg/L Roundup® were significantly smaller than those of the control group. The authors contextualized their findings by comparing them to the general US EPA environmental guideline limit for glyphosate and the California State’s specific EPA environmental guideline limit for glyphosate, which were 0.7 and 1.0 mg/L, respectively. These limits for glyphosate in surface waters are between the 0.45 and 1.35 mg/L concentrations Cuhra et al. used in their study [44]. They concluded that since D. magna showed complete reproductive failure and aborted all eggs in early-to-late stages of embryonic development when exposed to 1.35 mg/L suggests that the US EPA and the California State’s environmental guidelines may not be sufficiently restrictive to ensure viable populations of D. magna and other aquatic invertebrates.
Studies with rats seemed to suggest that the respiratory and hepatic systems as well as reproductive functions including sperm production or libido, and even foetal development can be altered by exposure to Roundup®. This may be attributed to the fact that xenobiotics in the aquatic ecosystem may lead to endocrine disruption at a reproductive and more specifically testicular level in young as well as adult mammals [45]. For example, different forms of testicular dysgenesis (abnormal development and growth of the testicles) have been linked to xenobiotics in aquatic ecosystems. These include decrease in sperm quantity and quality, increase in congenital malformations such as cryptorchidism (the absence of one or both testes from the scrotum) and hypospadias (abnormally placed urinary meatus), and preoccupying increase of testicular cancer incidences [46-49].
In a recent study, Prasad et al [50] investigated the genotoxic effects of glyphosate in the cells of Swiss Albino mice by measuring chromosomal aberrations (CAs) and micronuclei (MN) in bone marrow cells after exposure for 24, 48 and 72 h. Glyphosate treatment group mice were exposed by giving them a single dose of glyphosate intraperitoneally (i.p) at a concentration of 25 and 50 mg/kg b.wt. Simultaneously, positive control group mice were injected i.p. benzo(a)pyrene (100 mg/kg b.wt, once only), while control (vehicle) group mice were injected i.p. dimethyl sulfoxide (0.2 mL). Mice from all the groups were sacrificed at sampling times of 24, 48, and 72 h and their bone marrow analysed for cytogenetic and chromosomal damage. They reported that CAs and MN induction increased significantly in glyphosate treatment groups at both given dosages and time compared with the vehicle control group (P < 0.05). They also reported that glyphosate caused cytotoxic effects in the mice by significant decrease in mitotic index (MI). Based on their findings, Prasad et al [50] concluded that glyphosate is clastogenic and cytotoxic to mouse bone marrow.
In another study by Clair et al., mature rat fresh testicular cells were exposed to glyphosate and its formulation, Roundup®, from 1 to 10000 ppm [51]. This is the concentration range reported in some human urine and the environment, as well as in agricultural application levels. They found that Leydig cells got damaged from 1 to 48 h of Roundup® exposure, while other cells where damaged within 24-48 h; all mainly caused by necrosis. By contrast, glyphosate alone was toxic on Sertoli cells and later induced apoptosis at higher doses in germ cells and in Sertoli/germ cells co-cultures. At lower concentrations (i.e. 1 ppm), Roundup® and glyphosate were found to impact the endocrine system as they caused 35% decrease in testosterone, but only a high contamination appears to induce an acute rat testicular toxicity.
The effects of glyphosate on wildlife is not all that gloomy. At least, one study has reported this. Acacia et al. [52] studied the effects of glyphosate (as an active ingredient) and Roundup ® (as a formulation) on oyster gametes and embryos to find a possible link between genotoxicity and reproduction/developmental impairment. They wanted to explore the impact of chemical genotoxicity on population dynamics of oysters since glyphosate is frequently found in oyster production areas, among other herbicides. Considering that oyster’s gametes and embryos are in direct contact with the surrounding waters because its mode of reproduction is external, the presence of these agrochemicals does not only pose risk to oysters but also other aquatic organisms. In their study, Akcha et al. [52] exposed oyster spermatozoa and embryos to 0.5; 1.0; 1.5; 2.5; 5.0 μg active substance/L of both glyphosate and Roundup®. They reported that glyphosate and Roundup® had no effect on the oyster development at the concentrations tested. Their spermiotoxicity study also showed neither glyphosate nor Roundup® to be cytotoxic for oyster spermatozoa. It should be noted that although these findings by Akcha et al. [52] showed no negative effect on sperm function, the possible impact on fertilization rate and the consequences of the transmission of damaged DNA for oyster development and physiological performances was not investigated. More importantly, Akcha et al. [52] findings suggest that proper monitoring of usage of agrochemicals has the potential of minimising their toxic effects in the environment.
4. Toxicity of glyphosate and glyphosate-based herbicides to humans
Since the commercial introduction of glyphosate as a herbicide, its health effects have been studied intensely with the general conclusion that it is safe for humans [27, 42, 53]. However, recent published studies indicate that occupational exposure of humans to the herbicide is associated with increased cancer risks [54-55]. Also during the course of production of the herbicide, humans may come into dermal and/or inhalative contact with it. Although the shikimic pathway through which glyphosate disrupts plants biochemical activities is not found in animals, various studies seem to suggest the agrochemical may affect other pathways. The mitochondria and cytochrome P450 (CYP) pathways are thought to include possible candidate sites of action in animals, although there is no solid evidence to support such claims.
Mesnage et al. [56] investigated potential toxicity of 9 glyphosate-based formulations as well as technical grade glyphosate and polyethoxylated tallowamine POE-15 (the major adjuvant used in glyphosate-based formulations) to human cells, including hepatic (HepG2), embryonic (HEK293) and placental (JEG3) cell lines, after 24 h exposures [56]. They measured mitochondrial activities, membrane degradations and caspases 3/7 activities as endpoints. The authors reported that all formulations were more toxic than glyphosate, but POE-15 was found to be the most toxic against human cells, even if others were not excluded. The toxicity effect began with negative dose-dependent effects on cellular respiration and membrane integrity between 1 and 3 mg/L at environmental/occupational doses. They reported that POE-15 induced necrosis when its first micellisation process occurred, while glyphosate promoted endocrine disrupting effects after entering cells. The findings of Mesnage et al. challenged the establishment of guidance values such as the acceptable daily intake (ADI) of glyphosate since these are mostly based on a long term in vivo test of glyphosate alone [56]. The authors suggested that it is imperative to assess whole formulations of pesticides as mixtures with adjuvants that could change their toxicity in pesticide toxicity investigations.
Another study by Samsel and Seneff [57] asserted that glyphosate is minimally toxic to humans since residues are found in food stuff, including sugar, corn, soy and wheat. In their opinion, the disruption of cytochrome P450 (CYP) enzymes activities by glyphosate is an overlooked component of its toxicity to mammals. In a recent study, they reported that glyphosate interferences with cytochrome P450 (CYP) enzymes and acts synergistically to disrupt the biosynthesis of aromatic amino acids by gut bacteria, as well as impairment in serum sulphate transport. According to the authors, one of the many crucial functions of CYP enzymes is detoxification of xenobiotics. Therefore, by disrupting CYP enzymes activities, glyphosate enhances the damaging effects of other food borne chemical residues and environmental toxins. This adversely affects the body though the impact is subtle and manifests slowly over time as inflammation damages cellular systems throughout the body. Ultimately, these result in diseases and conditions such as gastrointestinal disorders, obesity, diabetes, heart disease, depression, autism, infertility, cancer and Alzheimer’s disease; mostly associated with a Western diet. Based on their study outcome, Samsel and Seneff described glyphosate as “textbook example” of exogenous semiotic entropy: the disruption of homeostasis by environmental toxins”.
Further recent studies have also suggested glyphosate to be an endocrine disrupting chemical (EDC), which has the potential to cause adverse health effects in humans [58]. In a study to evaluate the EDC properties of glyphosate in humans, Thongprakaisang et al [58] investigated the effects of technical grade glyphosate on estrogen receptors (ERs) mediated transcriptional activity and their expressions [58]. They reported that the proliferative concentrations of glyphosate, which caused the activation of estrogen response element (ERE) transcription activity, were 5-13 fold more than the control in T47D-KBluc cells. However, the activation was inhibited by an estrogen antagonist, ICI 182780, which implied that the estrogenic activity of glyphosate was mediated via ERs. The findings of this investigation suggest that low and environmentally relevant concentration of glyphosate can disrupt the hormonal systems of humans. Furthermore, the effects demonstrate that glyphosate is or could act as a “xenoestrogen” and may be capable of inducing EREs in a manner slightly weaker but functionally similar to Estradiol (E2), the most potent human estrogen.
In another study to investigate the xenobiotic toxicity of glyphosate, Gasnier et al. exposed the human liver HepG2 cells to four different formulations and to glyphosate, and measured the cytotoxicity, genotoxicity, anti-estrogenic (on estrogen receptors (ERα) and (ERβ)) and antiandrogenic effects (on androgen receptor (AR)), as well as checked androgen to estrogen conversion by aromatase activity and mRNA [59]. They reported that all parameters were disrupted at sub-agricultural concentrations with all formulations within 24 h, with the effects more dependent on the formulation than on the glyphosate concentration. They also stated that concentration levels above 0.5 mg/L of the most active formulation (R400) caused a human cell endocrine disruption on the androgen receptor in MDA-MB453-kb2 cells, while concentration levels above 2 mg/L inhibited transcriptional activities on both ERs on the HepG2. The authors also reported that concentration levels above 10 mg/L disrupted aromatase transcription and activity, while cytotoxic effects started at concentration levels above 10 mg/L and DNA damages (genotoxic effects) at 5 mg/L.
Richard et al also investigated the effects of glyphosate and Roundup® on human placental JEG3 cells within 18 h with concentrations lower than those found with agricultural use [60]. They stated that both chemical substances were toxic to the human placental JEG3 cells and this effect increases with concentration and time but Roundup® was found to be more toxic than glyphosate. They also tested the effects of both chemical substances at lower environmentally nontoxic concentrations on aromatase, the enzyme responsible for estrogen synthesis. They reported that Roundup® disrupted aromatase activity and mRNA levels and interacted with the active site of the purified enzyme, but the effects of glyphosate were facilitated by adjuvants in microsomes or in cell culture. Based on their findings, Richard® et al suggested that glyphosate and Roundup® can induce endocrine and toxic effects in humans and other mammals.
In a separate study to ascertain whether glyphosate exposure may cause DNA damage and cancer in humans, Koller et al. investigated exposure of workers via inhalation to technical glyphosate and Roundup UltraMax, glyphosate-based herbicide [61]. They reported that the cytotoxic and genotoxic properties of glyphosate in the workers’ buccal epithelial cell line (TR146) induced acute cytotoxic effects at concentration 40 mg/L after 20 min, which were due to membrane damage and impairment of mitochondrial functions. Similarly, they stated that Roundup UltraMax induced release of extracellular lactate dehydrogenase at concentrations 80 mg/L, which indicates membrane damage. In their study, Koller et al. showed that both glyphosate and Roundup UltraMax induced DNA migration in single-cell gel electrophoresis assays at concentrations 20 mg/L [61]. The authors again showed that the frequencies of micronuclei and nuclear buds were elevated after 20 min exposure to 10-20 mg/L, as well as increase of nuclear aberrations that reflected DNA damage. However, nucleoplasmatic bridges were only enhanced by glyphosate at the highest dose (20 mg/L). The authors concluded that their findings suggest that inhalation of glyphosate and glyphosate-based herbicides by humans may cause DNA damage in exposed individuals since they found genotoxic effects after short exposure to concentrations that correspond to a 450-fold dilution of spraying used in agriculture [61].
5. Conclusion
In this chapter, we discussed the perceived “friendly” nature of glyphosate and glyphosate-based herbicides and their apparent non-toxicity to both wildlife and humans. This notion has been in existence ever since glyphosate was first introduced as a commercial herbicide in the 1970s largely because the herbicidal activity of glyphosate targets the inhibition of the shikimate pathway in particular, which is only present in plants and microorganisms. Thus glyphosate is classified by the US EPA as a Group E carcinogen, and therefore is non-carcinogenic to humans. However, current investigations involving glyphosate exposure to wildlife and humans show adverse effects resulting from genotoxicity, cytotoxicity and reproductive toxicity. These have been reviewed with examples in this chapter. Furthermore, we have demonstrated that the herbicide has an endocrine impact at very low environmentally relevant concentrations. This review of glyphosate herbicides needs to be taken seriously since the use of glyphosate-contaminated plant products as dietary supplements may pose a risk of cancer in humans because of their potential additive estrogenicity. In addition, the extensive use of glyphosate-based herbicides in genetically modified glyphosate-resistant plants grown for food and feed should be of grave concern since they can be sources of genotoxicity, cytotoxicity, and reproductive toxicity in wildlife and humans.
Author details
Paul K. Mensah*, Carolyn G. Palmer and Oghenekaro N. Odume
*Address all correspondence to: [email protected]
Institute for Water Research, Rhodes University, Grahamstown, South Africa
References
[1] Jurado, A.S., Fernandes, M.A.S., Videira, R.A., Peixoto, F.P. and Vicente, J.A.F. (2011). Herbicides: The Face and the Reverse of the Coin. An in vitro Approach to the Toxicity of Herbicides in Non-Target Organisms, Herbicides and Environment, Andreas Kortekamp (Ed.), ISBN: 978-953-307-476-4, InTech. Accessed September 2013 at: http://www.intechopen.com/articles/show ... reverseof- the-coin-an-in-vitro-approach-to-the-toxicity-of-herbicides-i.
[2] PRB (Population Reference Bureau) (2012). 2012 World population data sheet, published by the Population Reference Bureau, 1875 Connecticut Ave., NW, Suite 520, Washington, DC 20009 USA. Accessed December 2012 at: http://www.prb.org/ pdf12/2012-population-data-sheet_eng.pdf.
[3] Schwarenbach, R.P., Escher, B.I., Fenner, K., Hoffstetter, T.B., Johnson, C.A., Von Gunten, U. and Wehrli, B. (2006). The challenge of micropollutants in aquatic systems. Science, 313, 1072-1077.
[4] Acquavella, J., Alexander, B., Mandel, J., Gustin, C., Baker, B., Chapman, P., Bleeke, M. (2004). Glyphosate biomonitoring for farmers and their families: results from the farm family exposure study. Environmental Health Perspective, 112, 321-326.
[5] LeBlanc, G.A. (2007). Crustacean endocrine toxicology: a review. Ecotoxicology, 16, 61-81.
[6] Correia, T.G., Narcizo, A.M., Bianchini, A. and Moreira, R.G. (2010). Aluminum as an endocrine disruptor in female Nile tilapia (Oreochromis niloticus). Comparative Biochemistry and Physiology, Part C, 151, 461-466.
[7] Pedersen, M., Halldorsson, T.I., Mathiesen, L., Mose, T., Brouwer, A., Hedegaard, M., Steffen, L., Kleinjans, J.C.S., Besselink, H. and Knudsen, L.E. (2010). Dioxin-like exposures and effects on estrogenic and androgenic exposures and micronuclei frequency in mother–newborn pairs. Environment International, 36, 344-351.
[8] Abel P.D. (2002). Water Pollution Biology (2nd edn.), Taylor and Francis Ltd, London.
[9] London, L., Dalvie, M.A. and Cairncross, E. (2005). Approaches for regulating water in South Africa for the presence of pesticides. Water SA, 31(1), 53-59.
[10] Mihaich, E.M., Friederich, U., Caspers, N., Hall, A.T., Klecka, G.M., Dimond, S.S., Staples, C.A., Ortego, L.S. and Hentges, S.G. (2009). Acute and chronic toxicity testing of bisphenol A with aquatic invertebrates and plants. Ecotoxicology and Environmental Safety, 72, 1392-1399.
[11] Benstead, R.S., Baynes, A., Casey, D., Routledge, E.J., Jobling, S. (2011). 17 β-Oestradiol may prolong reproduction in seasonally breeding freshwater gastropod molluscs. Aquatic Toxicology, 101, 326-334.
[12] Guilherme, S., Santos, M.A., Gaivão, I., Pacheco, M. (2014). Are DNA-damaging effects induced by herbicide formulations (Roundup® and Garlon®) in fish transient and reversible upon cessation of exposure? Aquatic Toxicology, 155, 213-221.
[13] Mensah, P.K., Palmer, C.G., Muller, W.J. (2014). Lethal and sublethal effects of pesticides on aquatic organisms: the case of a freshwater shrimp exposure to Roundup®. In: Larramendy, M.L. and Soloneski, S. (Eds), Pesticides: Toxic Aspects, InTech Publications, Rijeka, Croatia, pp. 163-185.
[14] Lipika, P. and Patra, A.K. (2006). Haemoatopoietic alterations induced by carbaryl in Clarias batrachus (LINN). Journal of applied Sciences and Environmental Management, 10 (3), 5-7.
[15] Boran, M., Altinok, I., Capkin, E., Karacam, H. and Bcer, V. (2007). Acute toxicity of carbaryl, methiocarb and carbosulfan to the rainbow trout (Oncorhynchus mykiss) and guppy (Poecilia reticulate). Turkish Journal of Veterinary and Animal Science, 31.
[16] Storrs, S. I. and Kiesiecker (2004). Survivorship patterns of larval amphibians exposed to low concentrations of atrazine. Environmental Health Perspectives, 112 (10), 1054-1057.
[17] Maharaj S. (2005). Modelling the Behaviour and Fate of Priority Pesticides in South Africa. MSc Thesis, Department of Earth Sciences, University of Western Cape, South Africa.
[18] Scholz, N.L., Incardona, J.P., Baldwin, D.H., Berejikan, B.A., Dittman, A.H., Feist, B.E., Jordan, C. (2003). Evaluating the sublethal impacts of current use pesticides on the environmental health of salmonids in Columbia River Basin. Bonneville Power Administration FY 2003 Provincial Project Review, 1-41
[19] Khan, M.Z., Tabassum, R., Naqvi, S.N.H., Shah, E.Z., Tabassum, F., Ahmad, I., Fatima, F., Khan, M.F. (2003). Effect of cypermethrin and permethrin on cholinester ase activity and protein contents in Rana tigrina (Amphibia), Turkey Journal of Zoology, 27, 243-246.
[20] Radosevich, S.R., Holt J.S., Ghersa C.M. (2007). Ecology of Weeds and Invasive Plants: Relationship to Agriculture and Natural Resource Management (3rd edn.), Wiley-Interscience, Hoboken, USA.
[21] Wilson, C. (2009). Aquatic toxicology notes: predicting the fate and effects of aquatic and ditchbank herbicides. Soil and Water Science Department, Institute of Food and Agricultural Sciences, University of Florida, Gainesville, USA.
[22] Pérez, G.L., Vera, M.S., Miranda, L. (2011). Effects of Herbicide Glyphosate and Glyphosate-Based Formulations on Aquatic Ecosystems. In: Andreas Kortekamp (Ed.), Herbicides and Environment. InTech Publications, Rijeka, Croatia.
[23] Mink, P.J., Mandel, J.S., Sceurman, B.K., Lundin, J.I. (2012). Epidemiologic studies of glyphosate and cancer: A review. Regulatory Toxicology and Pharmacology, 63, 440-452.
[24] Tu, M., Hurd, C., Randall, J.M. (2001).Weed Control Methods Handbook: Tools and Techniques for Use in Natural Areas, The Nature Conservancy. Accessed July 2009 at: http:// tncweeds.ucdavis.edu.
[25] Tsui, M.T.K., Chu, L.M. (2003). Aquatic toxicity of glyphosate-based formulations: comparison between different organisms and the effects of environmental factors. Chemosphere, 52, 1189-1197.
[26] Zhou, D-M., Wang, Y-J., Cang, L., Hao, X-Z., Luo, X-S. (2004). Adsorption and cosorption of cadmium and glyphosate on two soils with different characteristics. Chemosphere, 57, 1237-1244.
[27] WHO (World Health Organization) (1994). Environmental Health Criteria 159 – Glyphosate, International Programme on Chemical Safety. World Health Organization, Geneva, Switzerland.
[28] Miller, A., Gervais, J.A., Luukinen, B., Buhl, K., Stone, D. (2010). Glyphosate Technical Fact Sheet; National Pesticide Information Center, Oregon State University Extension Services. Accessed January 2012 at: http://npic.orst.edu/factsheets/glyphotech.html.
[29] Nordby, D.E., Hager, A.G. (2011). Herbicide formulations and calculations: active ingredient or acid equivalent, a Weed Fact sheet. Integrated Pest Management Handbook, University of Illinois, USA. Accessed 4 November 2011 at: http://ipm.illinois.edu/ weeds/aeai.pdf.
[30] Stenersen, J. (2004). Chemical Pesticides: Mode of Action and Toxicology. CRC Press, Boca Raton, Florida, USA, 296 pp.
[31] Schuette, J. (1998). Environmental fate of glyphosate, Environmental Monitoring & Pest Management, Department of Pesticide Regulation, Sacramento, CA 95824-5624, USA.
[32] Meyer, M.T., Loftin, K.A., Lee, E.A., Hinshaw, G.H., Dietze, J.E., Scribner, E.A. (2009). Determination of Glyphosate, its Degradation Product Aminomethylphosphonic Acid, and Glufosinate, in Water by Isotope Dilution and Online Solid-Phase Extraction and Liquid Chromatography/Tandem Mass Spectrometry. U.S. Geological Survey Techniques and Methods, book 5, chap. A10, 32p.
[33] Giesy, J.P., Dobson, S., Solomon, K.R. (2000). Ecotoxicological risk assessment for Roundup herbicide. Review of Environmental Contamination and Toxicology, 167, 35-120.
[34] Glusczak, L., Miron, D.S., Moraes, B.S., Simões, R.R., Schetinger, M.R.C., Morsch, V.M., Loro, V.L. (2007). Acute effects of glyphosate herbicide on metabolic and enzymatic parameters of silver catfish (Rhamdia quelen). Comparative Biochemistry and Physiology- Part C, 146, 519-524.
[35] El-Shebly, A.A., El-Kady, M.A.H. (2008). Effects of glyphosate herbicide on serum growth hormone (GH) levels and muscle protein content in Nile Tilapia (Oreochromis niloticus L.). Research Journal of Fisheries and Hydrobiology, 3(2), 84-88.
[36] vom Saal, F.S., Guillette, J.R. Jr., Myers, J.P., Swan, S. (2008). Endocrine disruptors: Effects in wildlife and laboratory animals. In: Encyclopaedia of Ecology, Jorgensen, S.E., Fath, B.D. (Eds). Pp 1261-1264.
[37] US EPA (1993). United States Environmental Protection Agency. Reregistration eligibility decision (RED): glyphosate. EPA 738-R-93-014. Office of Prevention, Pesticides, and Toxic Substances. Washington, DC: US EPA.
[38] Solomon, K., Anad, A., Carrasquilla, G., Cerdeira, A., Marshall, E., Sanin, L. (2007). Coca and poppy eradication in Colombia: Environmental and human health assessment of aerially applied glyphosate. Reviews of Environmental Contamination and Toxicology, 190, 43-125.
[39] EC (European Commission) (2002). Review report for the active substance glyphosate, Directive 6511/VI/99. January 21, 2002. Directorate E – Food Safety: plant health, animal health and welfare, international questions, E1-Plant health.
[40] WHO/FAO (World Health Organization/Food and Agriculture Organization of the United Nations) (2004). Pesticides Residues in Food, Report of the Joint Meeting of the FAO Panel of Experts on Pesticide Residues in Food and the Environment and the WHO Core Assessment Group on Pesticide Residues (JMPR). Rome, Italy, 20-29 September 2004. FAO Plant Production and Protection Paper 178. Rome, Italy.
[41] Shah, S.U. (2012). Importance of genotoxicity and S2A guidelines for genotoxicity testing for pharmaceuticals. IOSR Journal of Pharmacy and Biological Sciences, 1(2), 43-54.
[42] UNECE (United Nations Economic Commission for Europe) (2013). Globally Harmonized System of Classification and Labelling of Chemicals (GHS), (5th revised edn.), Part 3: Health Hazards. GHS (Rev.5). Accessed January 2015 at: http://www.unece.org/ru/ trans/danger/publi/ghs/ghs_rev05/05files_r.html.
[43] Vera-Candioti, J., Soloneski, S., Larramendy, M.L. (2013). Evaluation of the genotoxic and cytotoxic effects of glyphosate-based herbicides in the ten spotted live-bearer fish Cnesterodon decemmaculatus (Jenyns, 1842). Ecotoxicology and Environmental Safety, 89, 166-173.
[44] Cuhra, M., Traavik, T., Bøhn, T. (2013). Clone- and age-dependent toxicity of a glyphosate commercial formulation and its active ingredient in Daphnia magna. Ecotoxicology, 22, 251-262.
[45] Anway, M.D., Memon, M.A., Uzumcu, M., Skinner, M.K. (2006). Transgenerational effect of the endocrine disruptor vinclozolin on male spermatogenesis. Journal of Andrology, 27, 868-879.
[46] Carlsen, E., Giwercman, A., Keiding, N., Skakkebaek, N.E. (1992). Evidence for decreasing quality of semen during past 50 years. BMJ 305, 609-613.
[47] Auger, J., Kunstmann, J.M., Czyglik, F., Jouannet, P. (1995). Decline in semen quality among fertile men in Paris during the past 20 years. New England Journal of Medicine, 332, 281-285.
[48] Bergstrom, R., Adami, H.O., Mohner, M., Zatonski, W., Storm, H., Ekbom, A., Tretli, S., Teppo, L., Akre, O., Hakulinen, T. (1996). Increase in testicular cancer incidence in six European countries: a birth cohort phenomenon. Journal of the National Cancer Institute, 88, 727-733.
[49] Toppari, J., Virtanen, H.E., Main, K.M., Skakkebaek, N.E. (2010). Cryptorchidism and hypospadias as a sign of testicular dysgenesis syndrome (TDS): environmental connection. Birth Defects Res. A Clin. Mol. Teratol. 88, 910-919.
[50] Prasad, S., Srivastava, S., Singh, M., Shukla, Y. (2009). Clastogenic effects of glyphosate in bone marrow cells of Swiss albino mice. Journal of toxicology, 2009. 6 pages.
[51] Clair, É., Mesnage, R., Travert, C., Séralini, G.É. (2012). A glyphosate-based herbicide induces necrosis and apoptosis in mature rat testicular cells in vitro, and testosterone decrease at lower levels. Toxicology in vitro, 26(2), 269-279.
[52] Akcha, F., Spagnol, C. and Rouxel, J. (2012). Genotoxicity of diuron and glyphosate in oyster spermatozoa and embryos. Aquatic Toxicology, 106-107, 104-113.
[53] Williams, G.M., Kroes, R., Munro, I.C. (2000). Safety evaluation and risk assessment of the herbicide Roundup and its active ingredient, glyphosate, for humans. Regulatory Toxicology and Pharmacology, 31, 117-165.
[54] Eriksson, M., Hardell, L., Carlberg, M., Akerman, M. (2008). Pesticide exposure as risk factor for non-Hodgkin lymphoma including histopathological subgroup analysis. International Journal of Cancer, 123, 1657-1663.
[55] Bolognesi, C., Carrasquilla, G., Volpi, S., Solomon, K.R., Marshall, E.J. (2009). Biomonitoring of genotoxic risk in agricultural workers from five Colombian regions: association to occupational exposure to glyphosate. Journal of Toxicology and Environmental Health Part A, 72, 986-997.
[56] Mesnage, R., Bernay, B., Séralini, G.-E. (2013). Ethoxylated adjuvants of glyphosate-based herbicides are active principles of human cell toxicity. Toxicology, 313, 122-128.
[57] Samsel, A., Seneff, S. (2013). Glyphosate’s Suppression of Cytochrome P450 Enzymes and Amino Acid Biosynthesis by the Gut Microbiome: Pathways to Modern Diseases. Entropy, 15(4), 1416-1463.
[58] Thongprakaisang, S., Thiantanawat, A., Rangkadilok, N., Tawit Suriyo, T., Satayavivad, J. (2013). Glyphosate induces human breast cancer cells growth via estrogen receptors. Food and Chemical Toxicology, 59, 129-136.
[59] Gasnier, C., Dumont, C., Benachour, N., Clair, E., Chagnon, M-C., Séralini, G-E. (2009). Glyphosate-based herbicides are toxic and endocrine disruptors in human cell lines. Toxicology, 262, 184-191.
[60] Richard, S., Moslemi, S., Sipahutar, H., Benachour, N., Seralini, G.E. (2005). Differential effects of glyphosate and roundup on human placental cells and aromatase. Environmental health perspectives, 716-720.
[61] Koller, V.J., Furhacker, M., Nersesyan, A., Misik, M., Eisenbauer, M., Knasmueller, S. (2012). Cytotoxic and DNA-damaging properties of glyphosate and roundup in human- derived buccal epithelial cells. Archives of Toxicology, 86, 805-813.