By Mike Ludwig
t r u t h o u t
09 November 2010
NOTICE: THIS WORK MAY BE PROTECTED BY COPYRIGHT
YOU ARE REQUIRED TO READ THE COPYRIGHT NOTICE AT THIS LINK BEFORE YOU READ THE FOLLOWING WORK, THAT IS AVAILABLE SOLELY FOR PRIVATE STUDY, SCHOLARSHIP OR RESEARCH PURSUANT TO 17 U.S.C. SECTION 107 AND 108. IN THE EVENT THAT THE LIBRARY DETERMINES THAT UNLAWFUL COPYING OF THIS WORK HAS OCCURRED, THE LIBRARY HAS THE RIGHT TO BLOCK THE I.P. ADDRESS AT WHICH THE UNLAWFUL COPYING APPEARED TO HAVE OCCURRED. THANK YOU FOR RESPECTING THE RIGHTS OF COPYRIGHT OWNERS.

(Image: Jared Rodriguez / t r u t h o u t; Adapted: IRRI Images, Libertinus)
A delegation of politicians and community activists gathered on August 7 in La Leonesa, a small farm town in Argentina, to hear Dr. Andres Carrasco speak about a study linking a popular herbicide to birth defects in Argentina's agricultural areas.
But the presentation never happened. A mob of about 100 people attacked the delegation before they could reach the local school where the talk was to be held.
Dr. Carrasco and a colleague locked themselves in a car as the mob yelled threats and beat on the vehicle for two hours. One delegate was hit in the spine and has since suffered lower-body paralysis. Another person was treated for blows to the head. A former provincial human rights official was hit in the face and knocked unconscious.
Witnesses said the angry crowd had ties to local officials and agribusiness bosses, and police made little effort to stop the violence, according to human rights group Amnesty International.
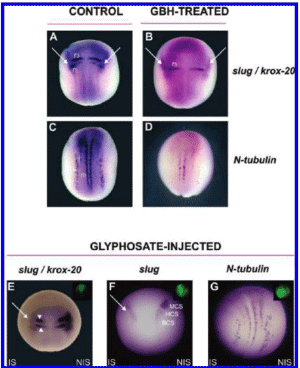
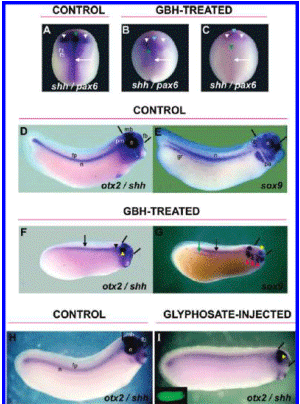
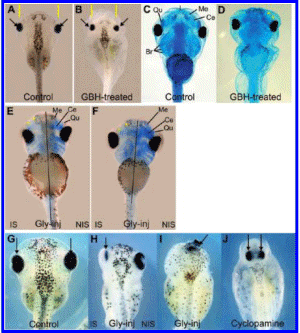
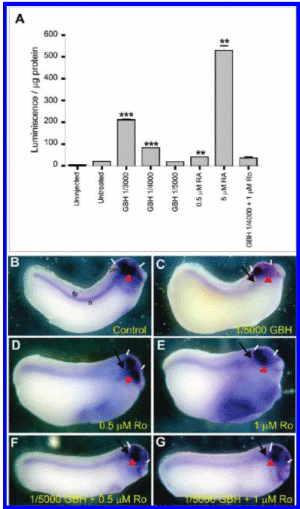
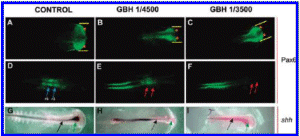
Carrasco is a lead embryologist at the University of Buenos Aires Medical School and the Argentinean national research council. His study, first released in 2009 and published in the United States this past summer, shows that glyphosate-based herbicides like Monsanto's popular Roundup formula caused deformations in chicken embryos that resembled the kind of birth defects being reported in areas like La Leonesa, where big agribusinesses depend on glyphosate to treat genetically engineered crops.
The deformations resulted from much lower doses of herbicide than those commonly found on crops, according to the study.
Biotech chemical giant Monsanto patented glyphosate under the trade name Roundup in the 1970's. The billion-dollar product is a main source of Monsanto's revenue and one of the most widely used herbicides in the world. One Monsanto blogger recently wrote that decades of success has made the Roundup brand name and glyphosate "interchangeable similar to the case of facial tissue and the brand name Kleenex."
Carrasco's report was largely ignored in the mainstream American media, but gained international attention among those opposed to genetically modified (GM) crops like Monstano's Roundup Ready crops, which are genetically engineered to tolerate the glyphosate-based herbicides.
The report is not the first to show that glyphosate herbicides like Roundup are more dangerous than government regulators and Monsanto have claimed, and Carrasco is not the first scientist to face intimidation after challenging the biotech industry, although he is the first to be threatened with violence.
Nevertheless, his report made an impact: journalists covered the results, environmentalists petitioned Argentina's high court to ban glyphosate and the government of the Argentinean province of Chaco began studying an eerie increase in birth defects and child cancer near the soy and rice fields sprayed with thousands of gallons of herbicide.
According to a spring 2010 report released by the Chaco government, an increase in birth defects and child cancer cases coincided with years of agricultural expansion and increased herbicide use in the province. The number of child cancer cases in La Leonesa, the small town where Carrasco and the other concerned citizens were attacked, has tripled from 2000 to 2009 and the number of birth defects in the province nearly quadrupled during that time, according to the report.
The report acknowledges that some local agribusinesses were unlawfully spraying herbicides too close to residential populations, but the Chaco study soon caught the attention of researchers across the world.
In September, an international coalition of scientists released a report citing the attack in La Leonesa and human tragedy in Chaco as proof that Roundup and genetically engineered soy crops are dangerous and unsustainable. The report provides a conclusive rebuttal to the industry's claims that spraying mutant crops with chemicals is the best way to feed the world. It's just another chapter in an information war that has raged for more than a decade, pitting independent scientists and embattled whistleblowers against the world's biggest biotech and petrochemical corporations.
Roundup and Monsanto
Monsanto has gained much of its international notoriety -- or infamy, depending on whom you talk to -- through its Roundup Ready line of crops that are genetically modified (GM) to be immune to the herbicide. To use the herbicide to combat weeds, farmers must buy patented Monsanto GM seeds with the genetic herbicide tolerant trait. Roundup herbicide is then sprayed to kill unwanted weeds, but the patented GM crops are spared.
The Roundup Ready crop system was first made available in 1996. Since 2000, the percentage of Roundup Ready corn grown in the United States has exploded from 7 to 70 percent and now 93 percent of the soybeans grown in the US are GM, according to the US Department of Agriculture (USDA).
Roundup accounts for about 40 percent of Monsanto's annual revenues and is sprayed on about 12 million acres of American farmland each year. In April, Monsanto announced the completion of a $200 million expansion of its glyphosate production facility in Louisiana.
Monsanto's Roundup Ready patent runs out in 2014, and the Justice Department began an antitrust investigation of Monsanto this year as its petrochemical competitors like DuPont clamor for a piece of the action. Monsanto has proven its tenacity in such disputes in the past; it forged new legal territory in the past decade, suing small farmers who saved Roundup Ready seeds or simply grew crops infected with GM traits after the patented Monsanto gene drifted and multiplied in their fields.
Superweeds
Monsanto's domination of domestic agriculture has had a startling side effect in the fields: the rise of new glyphosate resistant weeds commonly called "superweeds." Like the GM corn and soy, these weeds have bred themselves to tolerate Roundup and are invading farms across the country.
Monsanto shocked investors and environmentalists in October by announcing a new program that offers millions of dollars in rebates to farmers who combine Roundup with more herbicides manufactured by the company's competitors to combat the glyphosate-resistant weeds threatening GM crops across the country.
The mere presence of superweeds and the fact that Monsanto is now paying farmers to spray additional chemicals that are more toxic than Roundup, is evidence of a complete regulatory breakdown, according to watchdog group Center for Food Safety (CFS).
In his September 30 testimony to Congress on superweeds, CFS senior policy analyst William Freese said that the USDA regulates GM crops and the Environmental Protection Agency (EPA) regulates herbicides, but there is no regulation of the combined system.
"And it is the system -- the invariable use of glyphosate made possible and fostered by glyphosate-resistant seeds, for instance -- that is responsible for the growing epidemic of glyphosate-resistant (GR) weeds," Freese said in his testimony. "This is clearly demonstrated by the near complete absence of GR weeds for the first 20 plus years of glyphosate's use and the explosion of weed resistance in the decade since the widespread adoption of Roundup Ready crop systems."
Debate Gets Ugly
Glyphosate, the active ingredient in Roundup, has long been considered less toxic than other herbicides. The EPA considers glyphosate a noncarcinogen for humans and a chemical of relatively low toxicity.
Monsanto took the EPA's initial evaluation and ran with it, and in 1996, the state of New York filed a lawsuit against Monsanto over an advertising campaign that claimed Roundup to be as safe as table salt.
In recent years, teams of independent scientists like Carrasco's have come forward with studies showing that Roundup and glyphosate is more toxic than the regulators will admit. For years, Roundup critics charged that the "inert" ingredients like surfactants and solvents in Roundup and other glyphosate herbicides make the products more toxic to people and the environment.
Carrasco's report, on the other hand, showed that glyphosate itself caused malformations in embryos similar to those found in humans who live in agricultural areas dominated by genetically engineered crops. The report establishes that the toxic "inert" ingredients made it easier for the glyphosate to invade cells and cause damage.
But Carrasco is not the first scientist to identify this relationship between glyphosate and Roundup's "inert" ingredients.
Jeffrey Smith, GM critic and author of the books "Seeds of Deception" and "Genetic Roulette," told Truthout that many scientists have been verbally threatened and denied tenure for publishing studies critical of Roundup and GM crops.
"The attack [on Carrasco] is the latest in a series of attempts to silence those who have discovered problems with Roundup," Smith said.
Smith rattled off a list of scientists from Russia, Britain, the US, and beyond who have faced some kind of intimidation after going public with research on problems with GM foods and chemical products, including researcher Arpad Pusztai, who was famously relieved from his long-time position at a prominent Scottish research center in 1998 shortly after making public comments on potential problems with GM.
Smith is currently working with an international effort to support Gilles-Eric Seralini, a scientist at the University of Caen in France.
In 2009, Seralini and his team released a study showing that four different Roundup formulations diluted below suggested agricultural levels killed human placenta, umbilical chord and embryo cells.
"This clearly confirms that the [inert ingredients] in Roundup formulations are not inert," Seralini's team wrote. "Moreover, the proprietary mixtures available on the market could cause cell damage and even death around residual levels to be expected, especially in food and feed derived from [Roundup-treated] crops."
Carrasco cited Seralini's work in his groundbreaking study on glyphosate and birth defects.
Monsanto responded by calling Seralini's research "political" and argued that the conditions of the study did not reflect real life conditions. One Monsanto blogger even compared a key "inert" ingredient identified by Seralini's study to household soap.
Seralini and his team took on Monsanto again last year with a counteranalysis of lab data provided by Monsanto on the effects of three GM corn strains on lab rats. Seralini obtained the data after a German court ordered Monsanto to hand it over for review. Seralini's team discovered that the original study poorly constructed and the results reported by Monsanto were misleading.
Seralini had basically refuted Monsanto's ability to formally prove its GM products to be safe and that didn't sit well with his peers who supported the industry.
Pro-GM scientists in France, including Seralini's former colleague Marc Fellous of the French Association of Plant Biotechnology (AFBV), have since made public statements questioning Seralini's credibility and calling him a "merchant of fear," according to Seralini's supporters in the European scientific community.
Smith said that the intimidation of scientists conducting independent research, whether coming from the industry or its researchers, sends a dangerous message to other scientists.
"There is an attitude that, if you dare do research in the field, then you are threatening your work and credibility," Smith said.
As for Carrasco, the attack in La Leonesa did not keep him from speaking out. In September, just one month after being confronted by an angry mob, Carrasco was a featured speaker at the GMO-Free Europe conference.
Carrasco did not respond to a request for an interview.
Carrasco has his work cut out for him. On October 13, just days before initiating the plan to pay American farmers to use more herbicides, Monsanto announced that two more GM crops were approved in Argentina, according to a press release. Like the US, large Latin American countries like Argentina and Brazil are key growth markets for Monsanto.
This is the challenge facing Carrasco, Seralini, and others who use science to hold the biotech industry accountable for its push for control over the future of agriculture. Their stories show that taking on powerful financial interests of massive global corporations can be a difficult - and even dangerous - task: a war of information between those in search of profit and those in search of truth.