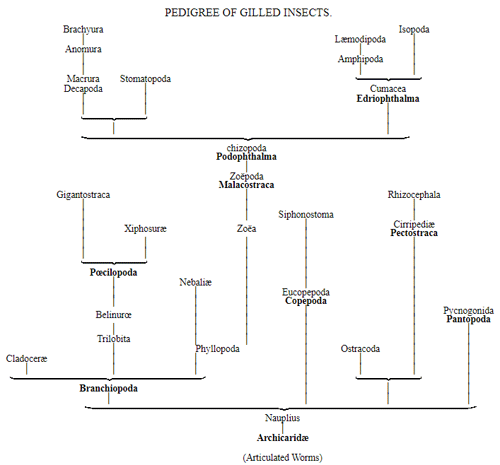
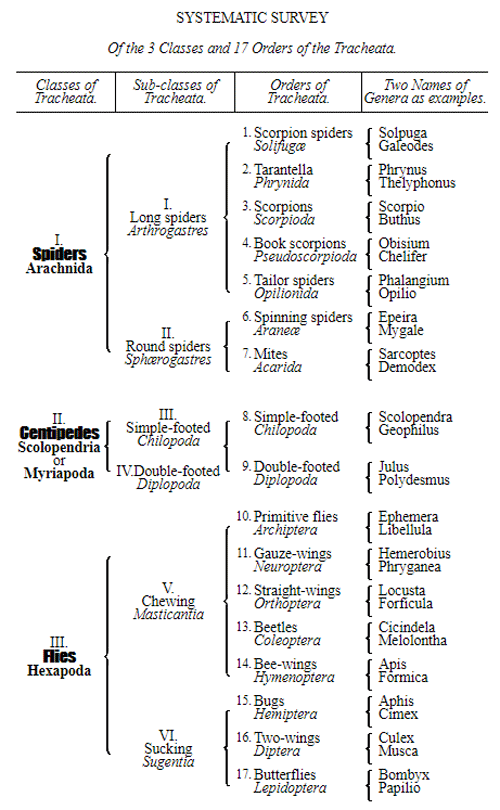
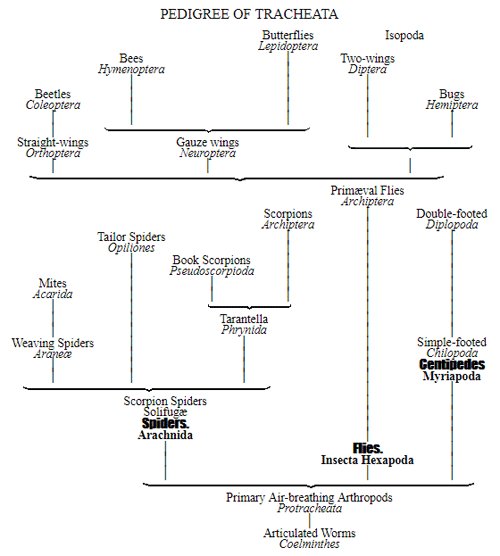
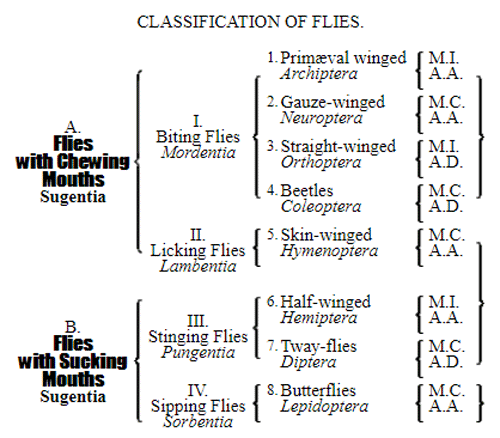
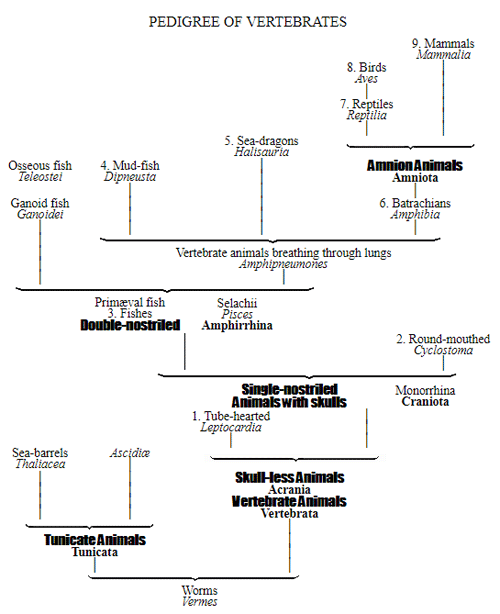
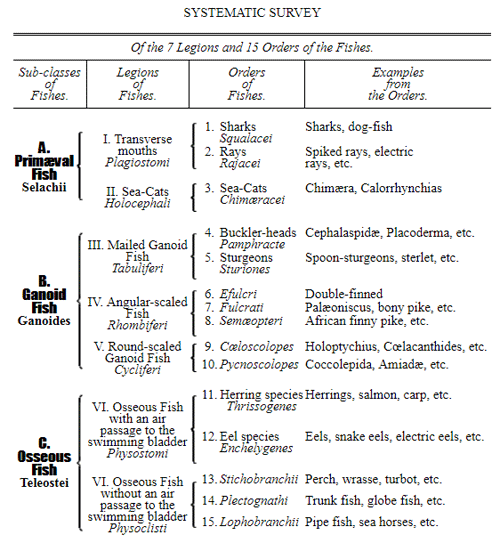
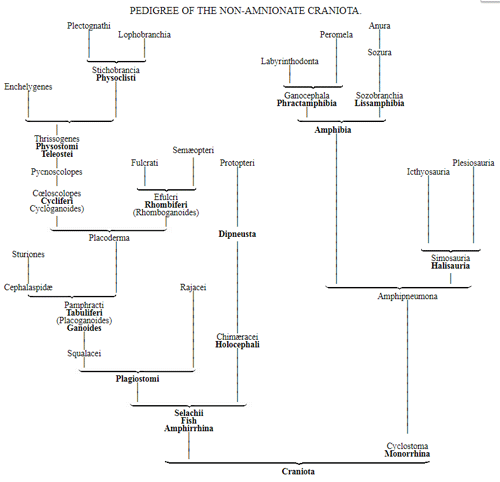
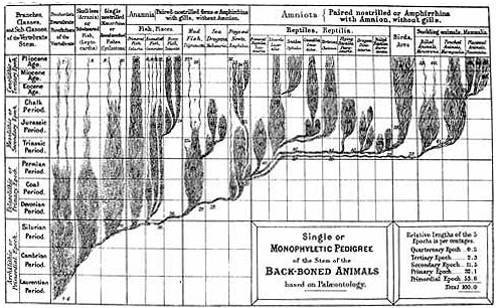
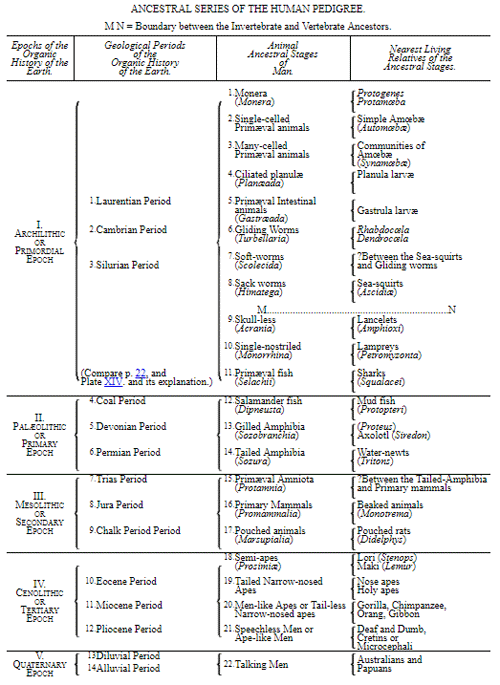
CHAPTER XVI. PEDIGREE AND HISTORY OF THE KINGDOM OF THE PROTISTA.
Special Mode of Carrying out the Theory of Descent in the Natural System of Organisms.—Construction of Pedigrees.—Descent of all Many-Celled from Single-Celled Organisms.—Descent of Cells from Monera.—Meaning of Organic Tribes, or Phyla.—Number of the Tribes in the Animal and Vegetable Kingdoms.—The Monophyletic Hypothesis of Descent, or the Hypothesis of one Common Progenitor, and the Polyphyletic Hypothesis of Descent, or the Hypothesis of Many Progenitors.—The Kingdom of Protista, or Primæval Beings.—Eight Classes of the Protista Kingdom.—Monera, Amœbæ, or Protoplastæ.—Whip-swimmers, or Flagellata.—Ciliated-balls, or Catallacta.—Labyrinth-streamers, or Labyrinthuleæ.—Flint-cells, or Diatomeæ.—Mucous-moulds, or Myxomycetes.—Root-footers (Rhizopoda).—Remarks on the General Natural History of the Protista: Their Vital Phenomena, Chemical Composition, and Formation (Individuality and Fundamental Form).—Phylogeny of the Protista Kingdom
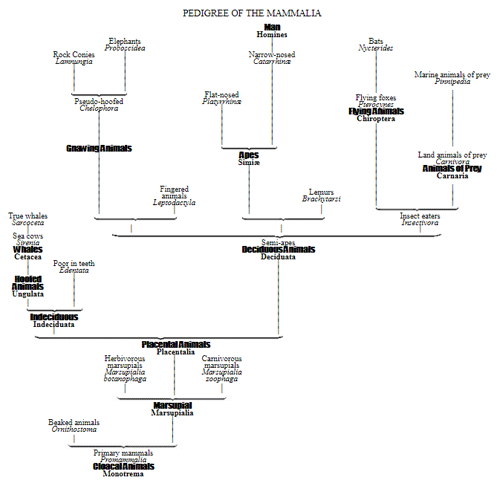
By a careful comparison of the individual and the palæontological development, as also by the comparative anatomy of organisms, by the comparative examination of their fully developed structural characteristics, we arrive at the knowledge of the degrees of their different structural relationships. By this, however, we at the same time obtain an insight into their true blood relationship, which, according to the Theory of Descent, is the real reason of the structural relationship. Hence by collecting, comparing, and 37 employing the empirical results of embryology, palæontology, and anatomy for supplementing each other, we arrive at an approximate knowledge of “the Natural System,” which, according to our views, is the pedigree of organisms. It is true that our human knowledge, in all things fragmentary, is especially so in this case, on account of the extreme incompleteness and defectiveness of the records of creation. However, we must not allow this to discourage us, or to deter us from undertaking this highest problem of biology. Let us rather see how far it may even now be possible, in spite of the imperfect state of our embryological, palæontological, and anatomical knowledge, to establish a probable scheme of the genealogical relationships of organisms.
Darwin in his book gives us no answer to these special questions of the Theory of Descent; at the conclusion he only expresses his conjecture “that animals have descended from at most only four or five progenitors, and plants from an equal or less number.” But as these few aboriginal forms still show traces of relationship, and as the animal and vegetable kingdoms are connected by intermediate transitional forms, he arrives afterwards at the opinion “that probably all the organic beings which have ever lived on the earth have descended from some one primordial form, into which life was first breathed by the Creator.” Like Darwin, all other adherents of the Theory of Descent have only treated it in a general way, and not made the attempt to carry it out specially, and to treat the “Natural System” actually as the pedigree of organisms. If, therefore, we venture upon this difficult undertaking, we must take up independent ground.
38 Four years ago I set up a number of hypothetical genealogies for the larger groups of organisms in the systematic introduction to my General History of Development (Gen. Morph. vol. ii.), and thereby, in fact, made the first attempt actually to construct the pedigrees of organisms in the manner required by the theory of development. I was quite conscious of the extreme difficulty of the task, and as I undertook it in spite of all discouraging obstacles, I claim no more than the merit of having made the first attempt and given a stimulus for other and better attempts. Probably most zoologists and botanists were but little satisfied with this beginning, and least so in reference to the special domain in which each one is specially at work. However, it is certainly in this case much easier to blame than to produce something better, and what best proves the immense difficulty of this infinitely complicated task is the fact that no naturalist has as yet supplied the place of my pedigrees by better ones. But, like all other scientific hypotheses which serve to explain facts, my genealogical hypotheses may claim to be taken into consideration until they are replaced by better ones.
I hope that this replacement will very soon take place; and I wish for nothing more than that my first attempt may induce very many naturalists to establish more accurate pedigrees for the individual groups, at least in the special domain of the animal and vegetable kingdom which happens to be well known to one or other of them. By numerous attempts of this kind our genealogical knowledge, in the course of time, will slowly advance and approach more towards perfection, although it can with certainty be foreseen that we shall never arrive at a 39 complete pedigree. We lack, and shall ever lack, the indispensable palæontological foundations. The most ancient records will ever remain sealed to us, for reasons which have been previously mentioned. The most ancient organisms which arose by spontaneous generation—the original parents of all subsequent organisms—must necessarily be supposed to have been Monera—simple, soft, albuminous lumps, without structure, without any definite forms, and entirely without any hard and formed parts. They and their next offspring were consequently not in any way capable of being preserved in a petrified condition. But we also lack, for reasons discussed in detail in the preceding chapter, by far the greater portion of the innumerable palæontological documents, which are really requisite for a safe reconstruction of the history of animal tribes, or phylogeny, and for the true knowledge of the pedigree of organisms. If we, therefore, in spite of this, venture to undertake their hypothetical construction, we must chiefly depend for guidance on the two other series of records which most essentially supplement the palæontological archives. These are ontogeny and comparative anatomy.
If thoughtfully and carefully we consult these most valuable records, we at once perceive what is exceedingly significant, namely, that by far the greater number of organisms, especially all higher animals and plants, are composed of a great number of cells, and that they originate out of an egg, and that this egg, in animals as well as in plants, is a single, perfectly simple cell—a little lump of albuminous constitution, in which another albuminous corpuscle, the cell-kernel, is enclosed. This cell containing its kernel grows and becomes enlarged. By division it forms an 40 accumulation of cells, and out of these, by division of labour (as has previously been described), there arise the numberless different forms which are presented to us in the fully developed animal and vegetable species. This immensely important process—which we may follow step by step, with our own eyes, any day in the embryological development of any animal or vegetable individual, and which as a rule is by no means considered with the reverence it deserves—informs us more surely and completely than all petrifactions could do as to the original palæontological development of all many-celled organisms, that is, of all higher animals and plants. For as ontogeny, or the embryological development of every single individual, is essentially only a recapitulation of phylogeny, or the palæontological development of its chain of ancestors, we may at once, with full assurance, draw the simple and important conclusion, that all many-celled animals and plants were originally derived from single-celled organisms. The primæval ancestors of man, as well as of all other animals, and of all plants composed of many cells, were simple cells living isolated. This invaluable secret of the organic pedigree is revealed to us with infallible certainty by the egg of animals, and by the true egg-cell of plants. When the opponents of the Theory of Descent assert it to be miraculous and inconceivable that an exceedingly complicated many-celled organism could, in the course of time, have proceeded from a simple single-celled organism, we at once reply that we may see this incredible miracle at any moment, and follow it with our own eyes. For the embryology of animals and plants visibly presents to our eyes in the shortest space of time the same process as that which has taken place in the 41 origin of the whole tribe during the course of enormous periods of time.
Upon the ground of embryological records, therefore, we can with full assurance maintain that all many-celled, as well as single-celled, organisms are originally descended from simple cells; connected with this, of course, is the conclusion that the most ancient root of the animal and vegetable kingdom was common to both. For the different primæval “original cells” out of which the few different main groups or tribes have developed, only acquired their differences after a time, and were descended from a common “primæval cell.” But where did those few “original cells,” or the one primæval cell, come from? For the answer to this fundamental genealogical question we must return to the theory of plastids and the hypothesis of spontaneous generation which we have already discussed (vol. i. p. 327).
As was then shown, we cannot imagine cells to have arisen by spontaneous generation, but only Monera, those primæval creatures of the simplest kind conceivable, like the still living Protamœbæ), Protomyxæ, etc. (vol. i. p. 1186, Fig. 1). Only such corpuscules of mucus without component parts—whose whole albuminous body is as homogeneous in itself as an inorganic crystal, but which nevertheless fulfills the two organic fundamental functions of nutrition and propagation—could have directly arisen out of inorganic matter by autogeny at the beginning (we may suppose) of the Laurentian period. While some Monera remained at the original simple stage of formation, others gradually developed into cells by the inner kernel of the albuminous mass becoming separated from the external cell-substance. In others, by differentiation of the outermost layer of the cell-substance, an external 42 covering (membrane, or skin) was formed round simple cytods (without kernel), as well as round naked cells (containing a kernel). By these two processes of separation in the simple primæval mucus of the Moneron body, by the formation of a kernel in the interior and a covering on the outer surface of the mass of plasma, there arose out of the original most simple cytods, or Monera, those four different species of plastids, or individuals, of the first order, from which, by differentiation and combination, all other organisms could afterwards develop themselves. (Compare vol. i. p. 347.)
The question now forces itself upon us, Are all organic cytods and cells, and consequently also those “original cells” which we previously considered to be the primary parents of the few great main groups of the animal and vegetable kingdoms, descended from a single original form of Moneron, or were there several different organic primary forms, each traceable to a peculiar independent species of Moneron which originated by spontaneous generation? In other words, Is the whole organic world of a common origin, or does it owe its origin to several acts of spontaneous generation? This fundamental question of genealogy seems at first sight to be of exceeding importance. But on a more accurate examination, we shall soon see that this is not the case, and that it is in reality a matter of very subordinate importance.
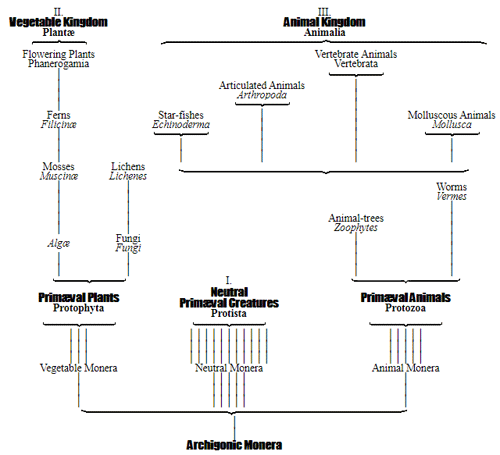
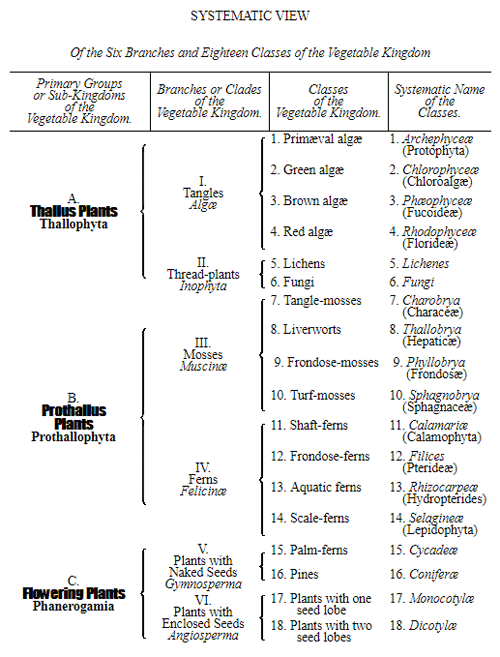
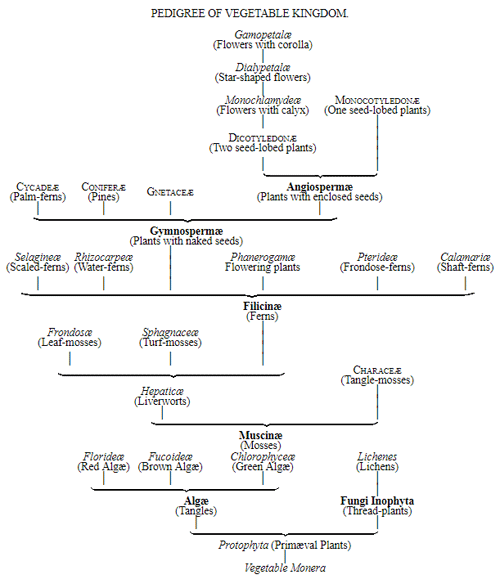
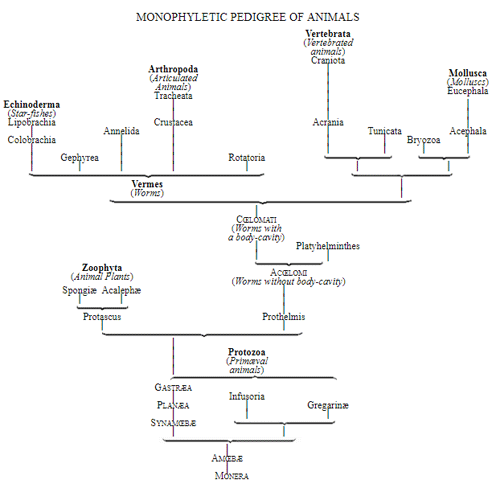
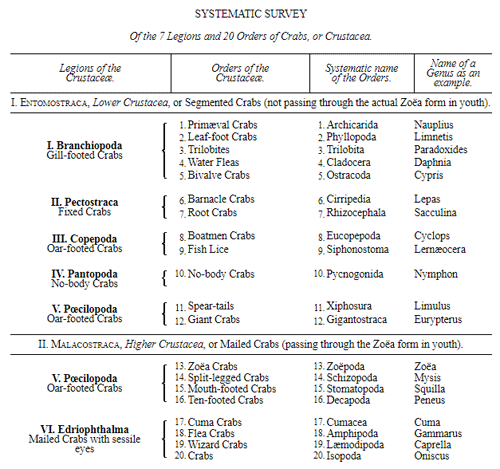
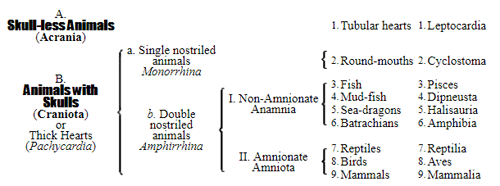
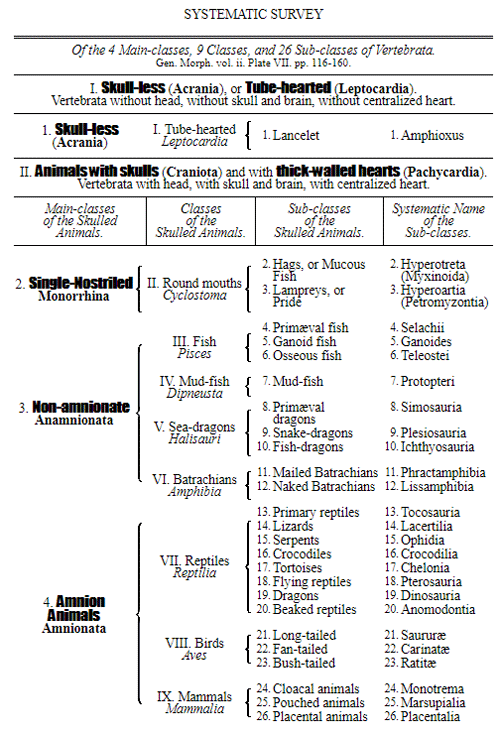
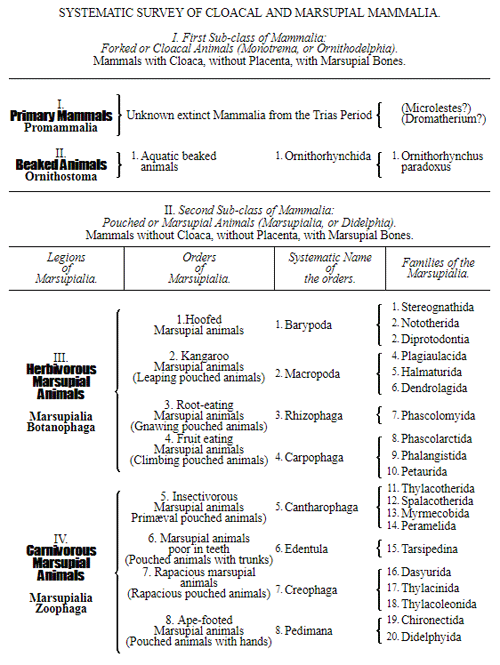
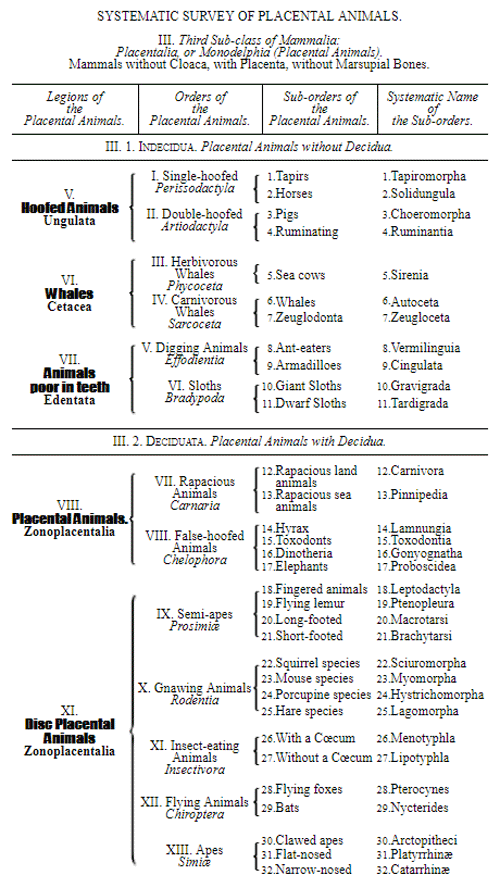
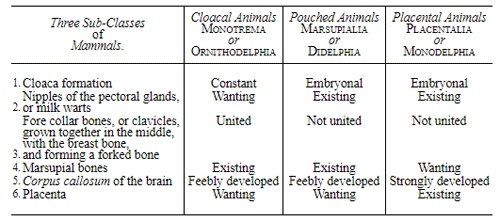
Let us now pass on to examine and clearly limit our conception of an organic tribe. By tribe, or phylum, we understand all those organisms of whose blood relationship and descent from a common primary form there can be no doubt, or whose relationship, at least, is most probable from anatomical reasons, as well as from reasons founded on historical 43 development. Our tribes, or phyla, according to this idea, essentially coincide with those few “great classes,” or “main classes,” of which Darwin also thinks that each contains only organisms related by blood, and of which, both in the animal and in the vegetable kingdoms, he only assumes either four or five. In the animal kingdom these tribes would essentially coincide with those four, five, or six main divisions which zoologists, since Bär and Cuvier, have distinguished as “main forms, general plans, branches, or sub-kingdoms” of the animal kingdom. (Compare vol. i. p. 53.) Bär and Cuvier distinguished only four of them, namely:—1. The vertebrate animals (Vertebrata); 2. The articulated animals (Articulata); 3. The molluscous animals (Mollusca); and 4. The radiated animals (Radiata). At present six are generally distinguished, since the tribe of the articulated animals is divided into two tribes, those possessing articulated feet (Arthropoda), and the worms (Vermes); and in like manner the tribe of radiated animals is subdivided into the two tribes of the star animals (Echinodermata) and the animal-plants (Zoophyta). Within each of these six tribes, all the included animals, in spite of great variety in external form and inner structure, nevertheless possess such numerous and important characteristics in common, that there can be no doubt of their blood relationship. The same applies also to the six great main classes which modern botany distinguishes in the vegetable kingdom, namely:—1. Flowering plants (Phanerogamia); 2. Ferns (Filicinæ); 3. Mosses (Muscinæ); 4. Lichens (Lichenes); 5. Fungi (Fungi); and 6. Water-weeds (Algæ). The last three groups, again, show such close relations to one another, that by the name of “Thallus plants” they may be contrasted with the three first main classes, and consequently 44 the number of phyla, or main groups, of the vegetable kingdom may be reduced to the number of four. Mosses and ferns may likewise be comprised as “Prothallus plants” (Prothallophyta), and thereby the number of plant tribes reduced to three—Flowering plants, Prothallus plants, and Thallus plants.
Very important facts in the anatomy and the history of development, both in the animal and vegetable kingdoms, support the supposition that even these few main classes or tribes are connected at their roots, that is, that the lowest and most ancient primary forms of all three are related by blood to one another. Nay, by a further examination we are obliged to go still a step further, and to agree with Darwin’s supposition, that even the two pedigrees of the animal and vegetable kingdom are connected at their lowest roots, and that the lowest and most ancient animals and plants are derived from a single common primary creature. According to our view, this common primæval organism can have been nothing but a Moneron which took its origin by spontaneous generation.
In the mean time we shall at all events be acting cautiously if we avoid this last step, and assume true blood relationship only within each tribe, or phylum, where it has been undeniably and surely established by facts in comparative anatomy, ontogeny, and phylogeny. But we may here point to the fact that two different fundamental forms of genealogical hypothesis are possible, and that all the different investigations of the Theory of Descent in relation to the origin of organic groups of forms will, in future, tend more and more in one or the other of these directions. The unitary, or monophyletic, hypothesis of descent will endeavour 45 to trace the first origin of all individual groups of organisms, as well as their totality, to a single common species of Moneron which originated by spontaneous generation (vol. i. p. 343). The multiple, or polyphyletic, hypothesis of descent, on the other hand, will assume that several different species of Monera have arisen by spontaneous generation, and that these gave rise to several different main classes (tribes, or phyla) (vol. i. p. 348). The apparently great contrast between these two hypotheses is in reality of very little importance. For both the monophyletic and the polyphyletic hypothesis of descent must necessarily go back to the Monera as the most ancient root of the one or of the many organic tribes. But as the whole body of a Moneron consists only of a simple, formless mass, without component particles, made up of a single albuminous combination of carbon, it follows that the differences of the different Monera can only be of a chemical nature, and can only consist in a different atomic composition of that mucous albuminous combination. But these subtle and complicated differences of mixture of the infinitely manifold combinations of albumen are not appreciable by the rude and imperfect means of human observation and are, consequently, at present of no further interest to the task we have in hand.
The question of the monophyletic or polyphyletic origin will constantly recur within each individual tribe, where the origin of a smaller or of a larger group is discussed. In the vegetable kingdom, for example, some botanists will be inclined to derive all flowering plants from a single form of fern, while others will prefer the idea that several different groups of Phanerogama have sprung from several different groups of ferns. In like manner, in the animal kingdom, 46 some zoologists will be more in favour of the supposition that all placental animals are derived from a single pouched animal; others will be more in favour of the opposite supposition, that several different groups of placental animals have proceeded from several different pouched animals. In regard to the human race itself, some will prefer to derive it from a single form of ape, while others will be more inclined to the idea that several different races of men have arisen, independently of one another, out of several different species of ape. Without here expressing our opinion in favour of either the one or the other conception, we must, nevertheless, remark that in general the monophyletic hypothesis of descent deserves to be preferred to the polyphyletic hypothesis of descent. In accordance with the chorological proposition of a single “centre of creation” or of a single primæval home for most species (which has already been discussed), we may be permitted to assume that the original form of every larger or smaller natural group only originated once in the course of time, and only in one part of the earth. We may safely assume this simple original root, that is, the monophyletic origin, in the case of all the more highly developed groups of the animal and vegetable kingdoms. (Compare vol. i. p. 353.) But it is very possible that the more complete Theory of Descent of the future will involve the polyphyletic origin of very many of the low and imperfect groups of the two organic kingdoms.
For these reasons I consider it best, in the mean time, to adopt the monophyletic hypothesis of descent both for the animal and for the vegetable kingdom. Accordingly, the above-mentioned six tribes, or phyla, of the animal kingdom 47 must be connected at their lowest root, and likewise the three or six main classes, or phyla, of the vegetable kingdom must be traced to a common and most ancient original form. How the connection of these tribes is to be conceived I shall explain in the succeeding chapters. But before proceeding to this, we must occupy ourselves with a very remarkable group of organisms, which cannot without artificial constraint be assigned either to the pedigree of the vegetable or to that of the animal kingdom. These interesting and important organisms are the primary creatures, or Protista.
All organisms which we comprise under the name of Protista show in their external form, in their inner structure, and in all their vital phenomena, such a remarkable mixture of animal and vegetable properties, that they cannot with perfect justice be assigned either to the animal or to the vegetable kingdom; and for more than twenty years an endless and fruitless dispute has been carried on as to whether they are to be assigned to this or that kingdom. Most of Protista are so small that they can scarcely, if at all, be perceived with the naked eye. Hence the majority of them have only become known during the last fifty years, since by the help of the improved and general use of the microscope these minute organisms have been more frequently observed and more accurately examined. However, no sooner were they better known than endless disputes arose about their real nature and their position in the natural system of organisms. Many of these doubtful primary creatures botanists defined as animals, and zoologists as plants; neither of the two would own them. Others, again, were declared by botanists to be plants, and by zoologists to be animals; each claimed them. These contradictions 48 are not altogether caused by our imperfect knowledge of the Protista, but in reality by their true nature. Indeed, most Protista present such a confused mixture of several animal and vegetable characteristics, that each investigator may arbitrarily assign them either to the animal or vegetable kingdom. Accordingly as he defines these two kingdoms, and as he looks upon this or that characteristic as determining the animal or vegetable nature, he will assign the individual classes of Protista in one case to the animal and in another to the vegetable kingdom. But this systematic difficulty has become an inextricable knot by the fact that all more recent investigations on the lowest organisms have completely effaced, or at least destroyed, the sharp boundary between the animal and vegetable kingdom which had hitherto existed, and to such a degree that its restoration is possible only by means of a completely artificial definition of the two kingdoms. But this definition could not be made so as to apply to many of the Protista.
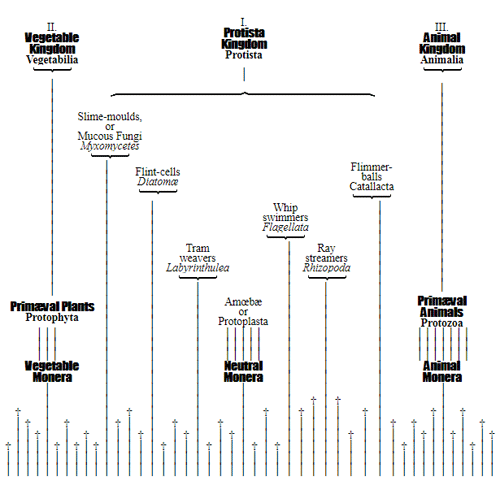
For this and other reasons it is, in the mean time, best to exclude the doubtful beings from the animal as well as from the vegetable kingdom, and to comprise them in a third organic kingdom standing midway between the two others. This intermediate kingdom I have established as the Kingdom of the Primary Creatures (Protista), when discussing general anatomy in the first volume of my General Morphology, pp. 191-238. In my Monograph of the Monera,(15) I have recently treated of this kingdom, having somewhat changed its limits, and given it a more accurate definition. Of independent classes of the kingdom Protista, we may at present distinguish the following:—
1. The still living Monera; 2. The Amœboidea, or Protoplasts; 3. The Whip-swimmers, or Flagellata; 4. The Flimmer-balls, or Catallacta; 5. The Tram-weavers, or Labyrinthuleæ; 6. The Flint-cells, or Diatomeæ; 7. The Slime-moulds, or Myxomycetes; 8. The Ray-streamers, or Rhizopoda.
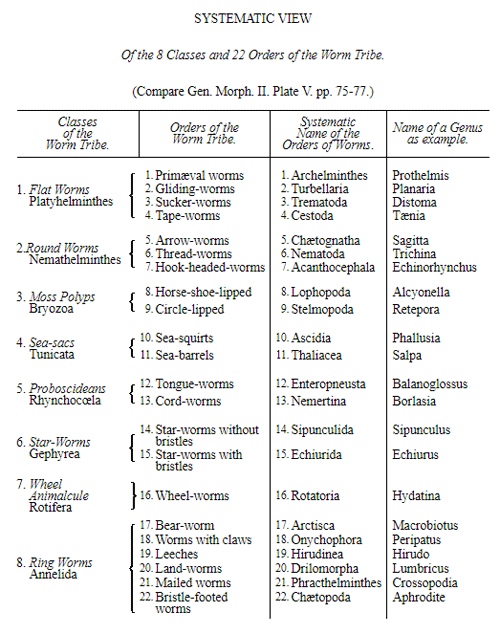
The most important groups at present distinguishable in these eight classes of Protista are named in the systematic table on p. 51. Probably the number of these Protista will be considerably increased in future days by the progressive investigations of the ontogeny of the simplest forms of life, which have only lately been carried on with any great zeal. With most of the classes named we have become intimately acquainted only during the last ten years. The exceedingly interesting Monera and Labyrinthuleæ, as also the Catallacta, were indeed discovered only a few years ago. It is probable also that very numerous groups of Protista have died out in earlier periods, without having left any fossil remains, owing to the very soft nature of their bodies. We might add to the Protista from the still living lowest groups of organisms—the Fungi; and in so doing should make a very large addition to its domain. Provisionally we shall leave them among plants, though many naturalists have separated them altogether from the vegetable kingdom.
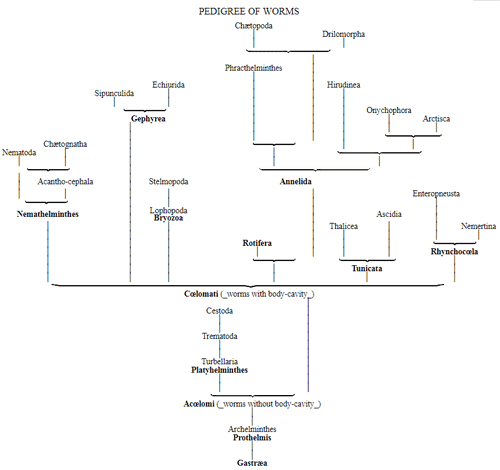
The pedigree of the kingdom Protista is still enveloped in the greatest obscurity. The peculiar combination of animal and vegetable properties, the indifferent and uncertain character of their relations of forms and vital phenomena, together with a number of several very peculiar features which separate most of the subordinate classes sharply from the others, at present baffle every attempt distinctly to make out their blood relationships with one 50 another, or with the lowest animals on the one hand, and with the lowest plants on the other hand. It is not improbable that the classes specified, and many other unknown classes of Protista, represent quite independent organic tribes, or phyla, each of which has independently developed from one, perhaps from various, Monera which have arisen by spontaneous generation. If we do not agree to this polyphyletic hypothesis of descent, and prefer the monophyletic hypothesis of the blood relationship of all organisms, we shall have to look upon the different classes of Protista as the lower small offshoots of the root, springing from the same simple Monera root, out of which arose the two mighty and many-branched pedigrees of the animal kingdom on the one hand, and of the vegetable kingdom on the other. (Compare pp. 74, 75.) Before I enter into this difficult question more accurately, it will be appropriate to premise something further as to the contents of the classes of Protista given on the next page, and their general natural history.
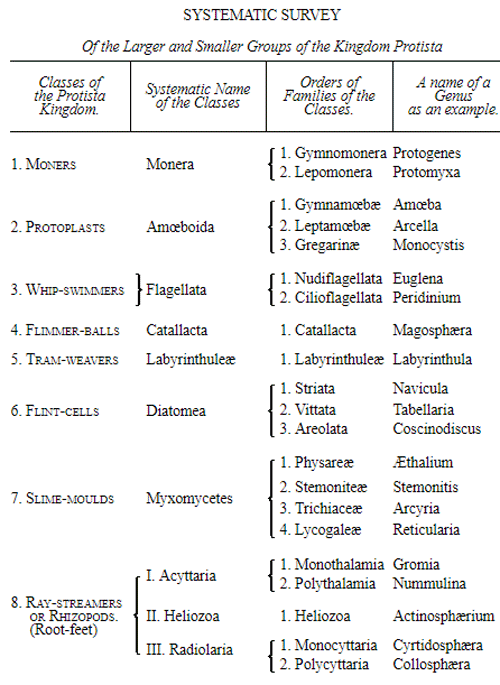
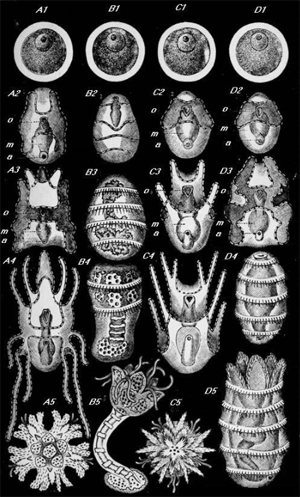
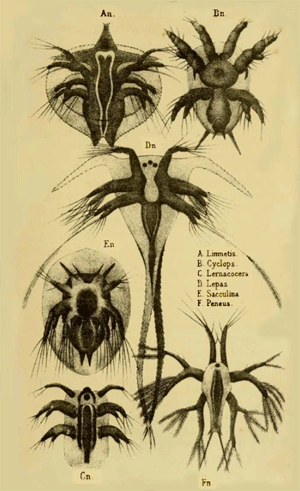
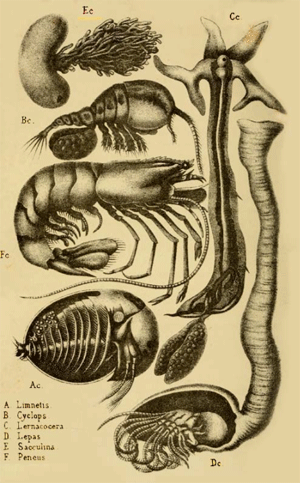
SYSTEMATIC SURVEY
Of the Larger and Smaller Groups of the Kingdom Protista
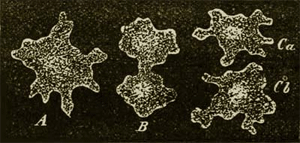
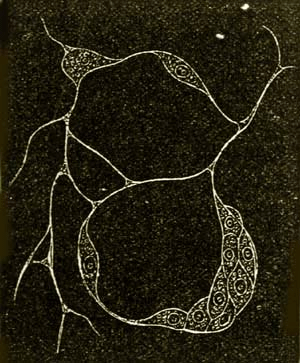
Fig. 8.—Protamœba primitiva, a fresh-water Moneron, much enlarged. A. The entire Moneron with its form-changing processes. B. It begins to divide itself into two halves. C. The division of the two halves is completed, and each now represents an independent individual.
It will perhaps seem strange that I should here again begin with the remarkable Monera as the first class of the Protista kingdom, as I of course look upon them as the most ancient primary forms of all organisms without exception. Still, what are we otherwise to do with the still living Monera? We know nothing of their palæontological origin, we know nothing of any of their relations to lower animals or plants, and we know nothing of their possible capability of developing into higher organisms. The simple and homogeneous little lump of slime or mucus which constitutes their entire body (Fig. 8) is the most ancient and original form of animal as well as of vegetable plastids. Hence it would evidently be just as arbitrary and unreasonable to assign them to the animal as it would be to assign them to the vegetable kingdom. In any case we shall for the present be acting more cautiously and critically if we comprise the still living Monera—whose number and distribution is probably very great—as a special and independent class, contrasting them with the other classes of the kingdom Protista, as well as with the animal kingdom. Morphologically considered, the Monera—on account of the perfect homogeneity of the albuminous substance of their bodies, on account of their utter want of heterogeneous particles—are more closely connected with anorgana than with organisms, and evidently form the transition between the inorganic and organic world of bodies, as is necessitated by the hypothesis of spontaneous generation. I have described and given illustrations of the forms and vital phenomena of the still living Monera (Protamœba, Protogenes, Protomyxa, etc.) in my Monograph of the Monera,(15) and have briefly mentioned the most important facts in the eighth chapter (vol. i. pp. 183-187). Therefore, only by way of a specimen, I here repeat the drawing of the fresh-water 53 Protamœba (Fig. 8). The history of the life of an orange-red Protomyxa adrantiaca, which I observed at Lanzerote, one of the Canary Islands, is given in Plate I. (see its explanation in the Appendix). Besides this, I here add a drawing of the form of Bathybius, that remarkable Moneron discovered by Huxley, which lives in the greatest depths of the sea in the shape of naked lumps of protoplasm and reticular mucus (vol. i. p. 344).
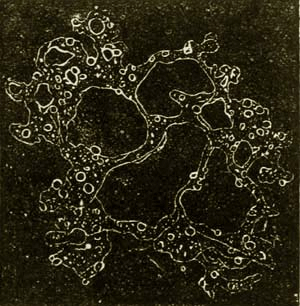
Fig. 9.—Bathybius Hæckelii, the “creature of primæval slime,” from the greatest depths of the sea. The figure, which is greatly magnified, only shows that form of the Bathybius which consists of a naked network of protoplasm, without the discoliths and cyatholiths which are found in other forms of the same Moneron, and which perhaps may be considered as the products of its secretion.
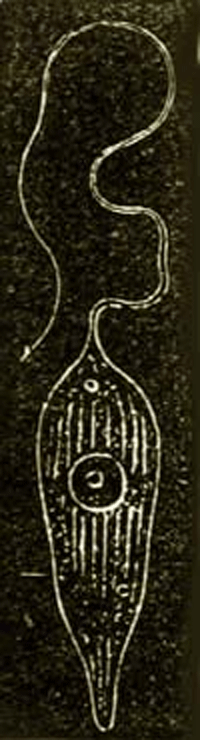
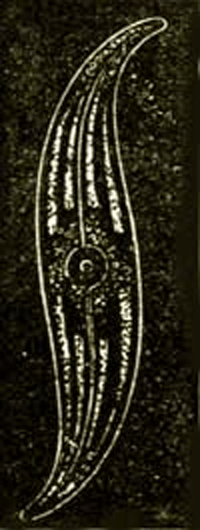
The Amœbæ of the present day, and the organisms most closely connected with them, Arcellidæ and Gregarinæ, which we here unite as a second class of Protista under the name of Amœboidea (Protoplasta), present no fewer genealogical difficulties than the Monera. These primary creatures are at present usually placed in the animal kingdom without its in reality being understood why. For simple naked cells—that is, shell-less plastids with a kernel—occur as well among real plants as real animals. The generative cells, for example, in many Algæ (spores and eggs) exist for a longer or shorter time in water in the 54 form of naked cells with a kernel, which cannot be distinguished at all from the naked eggs of many animals (for example, those of the Siphonophorous Medusæ). (Compare the figure of a naked egg of a bladder-wrack in Chapter xvii. p. 90.) In reality every naked simple cell, whether it proceeds from an animal or vegetable body, cannot be distinguished from an independent Amœba. For an Amœba is nothing but a simple primary cell, a naked little lump of cell-matter, or plasma, containing a kernel. The contractility of this plasma, which the free Amœba shows in stretching out and drawing in its changing processes, is a general vital property of the organic plasma of all animal as well as of all vegetable plastids. When a freely moving Amœba, which perpetually changes its form, passes into a state of rest, it draws itself together into the form of a globule, and surrounds itself with a secreted membrane. It can then be as little distinguished from an animal egg as from a simple globular vegetable cell (Fig. 10 A).
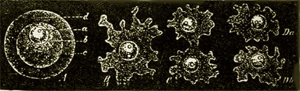
Fig. 10.—Amœba sphærococcus, greatly magnified. A fresh-water Amœba without a contractile vacuole. A. The enclosed Amœba in the state of a globular lump of plasma (c) enclosing a kernel and a kernel-speck (a). The simple cell is surrounded by a cyst, or cell membrane (d). B. The free Amœba, which has burst and left the cyst, or cell-membrane. C. It begins to divide by its kernel parting into two kernels, and the cell-substance between the two contracting. D. The division is completed, and the cell-substance has entirely separated into two bodies. (Da and Db)